: 30 Oct 2021, The Fellowship of the Ring, volume 88, no. 3
Since childhood, we have heard that nerve cells do not recover. And although the question of the possibility of the formation of new neurons in the adult brain is still open, there is already evidence that the process of neurogenesis in humans continues into old age. Any disturbances in the development of nerve cells can lead to serious, sometimes irreversible pathologies. One of these disorders is defects in the protective insulating sheath (myelin) of nerve cell processes, which can form in a person even before birth. They are almost impossible to diagnose using traditional imaging methods
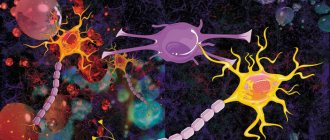
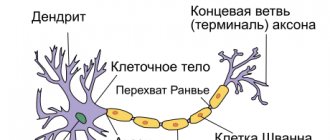
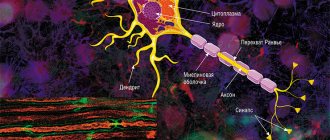
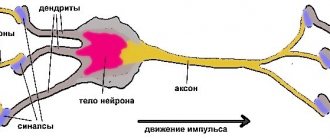
The average human brain contains about 100 billion neurons that receive, store, process and transmit information using electrical and chemical signals. Interaction between a neuron and other nerve cells and organs occurs through short ( dendrites)
) and long (
axon
) processes.
Each axon, like a wire, is covered with an insulating material - the myelin sheath
, which provides a higher speed of nerve impulses and protects nerve fibers from damage. In addition, this shell has a supporting function, and also, according to the latest data, serves as a kind of “refueling station” for the axon, which needs a large amount of energy.
All damage to the myelin sheath or defects that occur during its formation lead to serious, sometimes incurable diseases. Among them, the most famous is multiple sclerosis.
is a chronic autoimmune disease that primarily affects young people.
Myelin is also destroyed during strokes
, which occur not only in adults (primarily, as is commonly believed, in older people), but also in children, including the unborn.
Intrauterine stroke most often occurs after the 28th week of pregnancy, in children - a month after birth. Stroke in a fetus leads to the development of brain defects, and in children it can cause cerebral palsy
at an early age.
At the same time, today we judge the “quality” of myelination of the brain of a particular person only by indirect clinical symptoms or magnetic resonance imaging
(MRI), which can usually detect myelin defects at a late, often irreversible stage.
Neural insulation defects
Fetal brain development is a complex process in which rapid changes in the morphology and microstructure of nervous tissue occur. In some areas of the brain, the process of myelin formation begins as early as 18-20 weeks of pregnancy, and continues until approximately ten years of age.
Not everyone knows that myelin is many layers of cell membrane, “wound” many times around an axon. Myelin is formed by flat outgrowths of “service” glial cells, in which there is practically no cytoplasm. The myelin sheath is not continuous, but discrete, with gaps (nodes of Ranvier). Therefore, the axon has faster saltatory conduction: the speed of signal transmission along fibers with and without myelin can differ hundreds of times. As for the molecular composition of the “insulator”, it, like all cell membranes, consists mainly of lipids and proteins
It is myelination disorders that often underlie delays in the physical and mental development of a child, and also cause the formation of a number of neurological and psychiatric pathologies. In addition to diseases such as stroke, delays in fetal brain development with impaired myelination are sometimes observed in multiple pregnancies. At the same time, desynchronization in the development of the brain of twins is quite difficult to assess “by eye”.
But how to identify myelin defects during fetal development? Currently, obstetricians and gynecologists use only biometric indicators (for example, brain size), but these are highly variable and do not provide a complete picture. In pediatrics, even in the presence of obvious functional abnormalities in the child’s brain activity, traditional MRI or neurosonography
(ultrasound examinations of the brain of newborns) often do not show structural abnormalities.
Therefore, the search for accurate quantitative criteria for assessing myelin formation during pregnancy is an urgent task, which also needs to be solved using non-invasive diagnostic methods that have already been tested in obstetrics. Specialists from the Novosibirsk International Tomography Center SB RAS proposed using for these purposes a new method of quantitative neuroimaging, already adapted for prenatal ( prenatal)
) research.
Conduction of excitation in unmyelinated and myelinated nerve fibers
In soft nerve fibers, excitation spreads continuously along the entire membrane, from one excited area to another located nearby. In contrast, in myelinated fibers the action potential can propagate only spasmodically, “jumping” through sections of the fiber covered with an insulating myelin sheath. This type of conduction is called saltatory.
Direct electrophysiological studies carried out by Kato (1924) and then by Tasaki (1953) on single myelinated frog nerve fibers showed that action potentials in these fibers arise only in the nodes, and the areas between the nodes, covered with myelin, are practically inexcitable.
The density of sodium channels in the interceptions is very high: there are about 10,000 sodium channels per 1 μm2 of membrane, which is 200 times higher than their density in the membrane of the giant squid axon. A high density of sodium channels is the most important condition for saltatory conduction of excitation. Scheme in Fig. 46 allows us to understand how a nerve impulse “jumps” from one interception to another.
At rest, the outer surface of the excitable membrane of all nodes of Ranvier is positively charged. There is no potential difference between adjacent interceptions. At the moment of excitation, the surface of the membrane of interception A becomes charged electronegatively with respect to the surface of the membrane of the neighboring interception B. This leads to the emergence of a local electric current that flows through the interstitial fluid surrounding the fiber, the membrane and the axoplasm in the direction shown in Fig. 46 arrow. The current exiting through interception B excites it, causing the membrane to recharge. In interception A, the excitation still continues, and it temporarily becomes refractory. Therefore, interception B can only lead to a state of excitation of the next interception B, etc.
“Jumping” of the action potential across the interinterceptor region is possible only because the amplitude of the action potential in each interception is 5-6 times higher than the threshold value required to excite the neighboring interception. Under certain conditions, an action potential can “jump” not only through one, but also through two inter-intercept sections. This is observed, in particular, if the excitability of the neighboring interception is reduced by some pharmacological agent, for example novocaine, cocaine, etc.
| V.V. |
The time required to transfer excitation from one interception to another is approximately the same for fibers of different diameters (at a temperature of 24 °C it is about 0.07 ms). The length of the inter-intercept sections, as noted, is proportional to the diameter of the unwound fiber. It follows that in myelinated
| 2. | |
| 3 d'-L | |
| 4 D | |
| ■J | \ |
| '.L | |
| ^———— | |
| Shm | 1SHSHSH |
Rice. 47. Responses of the frog sciatic nerve to electrical stimuli of increasing strength.
The abduction of action potentials is biphasic. An irritation artifact is visible at the beginning of each recording. The current increases gradually from I to 6. The time stamp is 1 ms.
Rice. 46. Saltatory propagation of excitation in the mnogo nerve fiber from interception to interception.
The arrows show the direction of the current arising between the excited (A) and adjacent quiescent (B) interception.
fibers, the speed of nerve impulse conduction is approximately proportional to their diameter. In this respect, myelinated fibers differ from non-myelinated fibers, in which the conduction velocity is proportional not to the diameter, but to the square root of its value.
The conduction of excitation along a myelinated nerve fiber is often compared to the transmission of signals along an electrical cable with repeating generators (for example, the transatlantic cable). Indeed, the sections of nerve fiber between the interceptions are similar in their electrical properties to a cable immersed in a liquid with high electrical conductivity. The internal conductor is the axoplasm, the external conductor is the intercellular fluid, and the insulator is the fatty myelin sheath. The impulse passing between the interceptions is an electrical current impulse. Ranvier intercepts play the role of relay generators, i.e., intermediate amplification stations of the communication line. When transmitting a signal, each subsequent interception is excited by a pulse generated by the previous one, generates a new pulse and transmits it along the fiber. Since the resistance of the inner conductor per unit length is very high (106 times greater than that of copper wire of the same diameter), the repeater generators must be located close to each other, otherwise the pulse will die out.
The assumption about the spasmodic propagation of excitation in nerve fibers was first expressed by B. F. Verigo (1899). This method of conduction has a number of advantages compared to continuous conduction in non-pulp fibers: firstly, by “jumping” over relatively large sections of the fiber, excitation can spread at a much higher speed than with continuous conduction: along a non-pulp fiber of the same diameter; secondly, abrupt propagation is energetically more economical, since not the entire membrane comes into a state of activity, but only its small sections in the interception area, having a width of less than 1 μm. The losses of ions (per unit fiber length) accompanying the occurrence of an action potential in such limited areas of the membrane are very small, and therefore the energy costs for the operation of the sodium-potassium pump, necessary to restore the altered ionic ratios between the internal contents of the nervous system, are also small. fibers and tissue fluid.
the composite nature of the action potential of the nerve trunk and
classification of nerve fibers
The amplitude of electrical impulses removed from the entire nerve trunk depends on the strength of the applied stimulus. A weak stimulus corresponds to
| j—1 IL |
| 100 — 75 -60 — |
| i III i III |
With
25 50 75 100 125 150 175 200 225 250 275 300
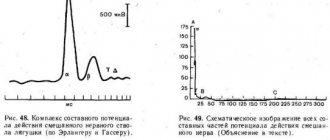
Rice. 48. Complex of the compound action potential of the mixed nerve trunk of the frog (according to Erlanger and Gasser).
Rice. 49. Schematic representation of all components of the action potential of a mixed nerve (Explanation in the text).
small answer; as stimulation intensifies, the amplitude of the potential increases, reaching a maximum value, and then remains constant, despite a further increase in the strength of the stimulus (Fig. 47). This is explained by the fact that the electrical response of the entire nerve trunk is an algebraic sum of the action potentials of its individual fibers. In each fiber, the amplitude of the action potential does not depend on the strength of stimulation in accordance with the “all or nothing” law. The stimulation thresholds of individual fibers differ from each other. With weak stimulus strength, excitation occurs in the most excitable superficial nerve fibers. Increasing the stimulus leads to an increase in the number of excited fibers, so the total response to stimulation increases until all fibers are involved in the reaction.
This picture can be observed if the discharge electrodes are placed on the nerve near the stimulating electrodes. As the distance between these two pairs of electrodes increases, the total action potential begins to be divided into several individual oscillations, which become most clearly expressed when the output electrodes move 10-15 cm from the site of stimulation (Fig. 48). This phenomenon was first studied in detail by Erlanger and Gasser (1937). They showed that the reason for the division of the total action potential into components is the unequal speed of excitation along different fibers, as a result of which nerve impulses arrive at the output electrodes through these fibers non-simultaneously.
At present, nerve fibers, based on the speed of excitation, the duration of various phases of the action potential and structure, are usually divided into three main types, designated by the letters A, B and C (Fig. 49).
Type A fibers are divided into four subgroups: oc, (5, y, b. They are covered with a myelin sheath. The thickest of them are a-fibers (Aa), in warm-blooded animals and humans they have a diameter of 12-22 microns and are characterized by a significant conduction speed excitation - 70-120 m/s. Such fibers conduct excitation from the motor nerve centers of the spinal cord to the skeletal muscles (motor fibers) and from certain muscle receptors to the corresponding nerve centers. The peak action potential of the Aa fiber in warm-blooded animals lasts 0.4-0. 5 ms After its end, a trace depolarization develops, which lasts 15-20 ms and turns into a trace hyperpolarization lasting about 40-60 ms.
Three other groups of type A fibers: Ap, Au and A5 - have a smaller diameter, lower conduction velocity and a longer action potential. These are predominantly sensory fibers that conduct excitation from various receptors (tactile, some pain, temperature and receptors of internal organs) to the central nervous system.
The only exceptions are y-fibers, a significant part of which conduct excitation in a centrifugal direction from the cells of the spinal cord to the so-called intrafusal muscle fibers that are part of the muscle receptors - muscle spindles (see Fig. 51, A).
Type B fibers include myelinated, predominantly preganglionic, fibers of the autonomic nervous system. The speed of excitation in these fibers in warm-blooded animals is 3-18 m/s. The duration of the action potential of type B fibers (1-2 ms) is approximately 3 times longer than the duration of the action potential of type A fibers. A distinctive feature of these fibers is that they do not exhibit a trace depolarization phase: the descending leg of the peak directly turns into a trace hyperpolarization, which in some cases it lasts more than 100 ms.
Type C fibers are non-mellose nerve fibers with a very small diameter (approximately 1 µm). The excitation speed in these fibers is no more than 3 m/s. Most type C fibers are postganglionic fibers of the sympathetic nervous system.
Type C fibers also include those nerve fibers that are involved in conducting excitation from pain receptors and some receptors of cold, heat and pressure.
Action potentials of fibers of this type are characterized by the longest duration (2 ms in warm-blooded animals). They have a long phase of trace depolarization (50-80 ms), accompanied by an even longer (300-1000 ms) trace hyperpolarization. All these data are summarized in table. 3.
| Table 3 ⇐ Previous3Next ⇒ Recommended pages: |
On a regular tomograph
Any pathology of the fetal brain that doctors suspect during an ultrasound examination of a pregnant woman is usually an indication for an MRI; Similar studies have been carried out at the ITC SB RAS for more than ten years. MRI results can confirm, clarify, refute, or even change the preliminary diagnosis and, accordingly, pregnancy management tactics.
The fact is that the amount of myelin and the size of individual brain structures in the embryo are so small that any measurements are very complex and time-consuming. In addition, the fetus is constantly moving, which makes it very difficult to obtain high-quality images and reliable quantitative data. Therefore, we need technology that allows us to obtain images quickly and with high resolution even on small objects.
This is exactly what the method for fast mapping of the macromolecular proton fraction
(MPF) is a biophysical parameter that describes the proportion of protons in tissue macromolecules involved in the formation of the MRI signal, whereas the signal source is usually protons contained in water (Yarnykh, 2012; Yarnykh et
al
., 2015).
The method is based on a specialized procedure for mathematical processing of MRI images, which makes it possible to isolate signal components associated with the MPF of cell membranes. And in the brain of humans and animals, the main part of them is contained in myelin. MPF maps are reconstructed based on initial data, which can be obtained on almost any clinical tomograph.
To reconstruct the MPF maps, four source images obtained by various traditional MRI methods are used. The correctness of this approach was confirmed by the results of its testing on laboratory animals at Tomsk State University: in mice that were injected with a solution that causes myelin destruction, the results of MPF mapping coincided with the data of histological examination of tissues (Khodanovich et al
., 2017).