Classification of nerve fibers
| The amplitude of electrical impulses removed from the entire nerve trunk depends on the strength of the applied stimulation. A small action potential corresponds to stimulation of weak strength, but as stimulation increases, the amplitude of the peak increases, reaches a maximum value and then remains constant, despite a further increase in the strength of the stimulus (Fig. 156). Rice. 156. Responses of the sciatic nerve of the frog to electrical stimuli of increasing strength (according to J. Buresh). The abduction of action potentials is biphasic. At the beginning of each recording there is an irritation artifact. The current increases gradually from 1 to 6. Time stamp 1 ms. This is explained by the fact that the action potential removed from the entire nerve trunk is an algebraic sum of the potentials of its individual nerve fibers . In each fiber, the amplitude of the action potential does not depend on the strength of stimulation in accordance with the “all or nothing” law. The stimulation thresholds of individual fibers differ from each other. With weak stimulus strength, excitation occurs in the most excitable superficial nerve fibers. Increasing the stimulus leads to an increase in the number of excited fibers, so the total peak increases until all fibers are involved in the response. This picture can be observed if the discharge electrodes are placed on the nerve near the stimulating electrodes. As the distance between these two pairs of electrodes increases, the total action potential begins to be divided into several individual oscillations, which become most clearly expressed when the output electrodes move 10-15 cm from the site of stimulation (Fig. 157). |
| This phenomenon was first studied in detail by Erlanger and Gasser (1937). They showed that the reason for the division of the total action potential into components is the unequal speed of excitation along different fibers, as a result of which nerve impulses arrive at the output electrodes through these fibers non-simultaneously. Rice. 157. Complex of the compound action potential of the mixed nerve trunk of the frog (according to Erlanger and Gasser). |
| A detailed analysis showed that there is an approximately proportional relationship between the speed of impulse conduction and the diameter of the nerve fiber: nerve fibers conduct faster the thicker they are. Currently, it is customary to divide nerve fibers according to the speed of excitation, the duration of various phases of the action potential and structure into three main types, designated by the letters A, B and C (Fig. 158). Rice. 158. Schematic representation of all components of the action potential of a mixed nerve (according to Erlanger and Gasser). |
Classification of nerve fibers
Type A nerve fibers are divided into four subgroups: α, β, γ and δ. They are covered with a myelin sheath. The thickest of them - the so-called alpha fibers (Aα) - in warm-blooded animals and humans have a diameter of 12-22 microns and are characterized by a significant speed of excitation: 70-120 m/sec. Such fibers conduct excitation from the motor nerve centers of the spinal cord to the skeletal muscles (“motor” fibers) and from muscle receptors to the corresponding nerve centers. The action potential of Aα fibers in warm-blooded animals lasts 0.5 ms. After its completion, a trace negative potential develops, which lasts 15-20 ms and turns into a trace positive potential lasting about 40-60 ms.
Three other groups of type A fibers: beta (Aβ)-, gamma (Aγ)- and delta fibers have a smaller diameter - from 2 to 12 microns, a lower conduction velocity and a longer action potential. These are predominantly sensory fibers that conduct excitation from various receptors (tactile, some pain, temperature and receptors of internal organs) into the central nervous system. The only exceptions are gamma fibers, a significant part of which conduct excitation in a centrifugal direction - from the cells of the spinal cord to the so-called intrafusal muscle fibers of which sensitive nerve endings - muscle spindles - are located.
Type B fibers include mpelin, predominantly preganglionic fibers of the autonomic nervous system. The speed of excitation in these fibers in warm-blooded animals is 3-14 m/sec. The duration of the action potential of type B fibers is approximately 2 times longer than the duration of the action potential of type A fibers. On average, it is 1.2 ms. A distinctive feature of these fibers is that they do not exhibit a trace negative potential: the repolarization phase directly transforms into a trace positive potential, which has a long duration - 100-300 ms.
Type C fibers include soft nerve fibers of very small diameter (about 0.5 µm). The speed of excitation in these fibers is 2-6 m/sec. Most C fibers are postganglionic fibers of the sympathetic nervous system.
C-fibers also include those nerve fibers that are involved in conducting excitation from pain receptors and some receptors of cold, heat and pressure. Action potentials of C-fibers are characterized by the longest duration (2 ms in warm-blooded animals). They have a long trace negative potential (50-80 ms), accompanied by an even longer trace positivity (300-1000 ms).
All this data is summarized in the table.
Properties of various nerve fibers of warm-blooded animals
| Fiber type | Fiber diameter (μ) | Conduction speed (m/sec) | Action potential duration (ms) | Duration of negative trace potential (ms) | Duration of positive trace potential (ms) | Function |
| A (α) | 12-22 | 70-120 | 0,4-0,5 | 12-20 | 40-60 | Motor fibers of skeletal muscles, afferent fibers of muscle receptors |
| A(β) | 8-12 | 40-70 | 0,4-0,6 | — | — | Afferent fibers from touch receptors |
| A (γ) | 4-8 | 15-40 | 0,5-0,7 | — | — | Afferent fibers from touch and pressure receptors, efferent fibers to muscle spindles |
| A (Δ) | 1-4 | 5-15 | 0,6-1,0 | — | — | Afferent fibers from some receptors of heat, pressure, pain |
| IN | 1-3 | 3-14 | 1-2 | Absent | 100-300 | Preganglionic autonomic fibers |
| WITH | 0,5-1,0 | 0,5-2 | 2,0 | 50-80 | 300-1000 | Preganglionic autonomic fibers, afferent fibers from some pain, pressure, and heat receptors |
Normal physiology: lecture notes
2. Mechanisms for conducting excitation along the nerve fiber. Laws for the conduction of excitation along nerve fibers
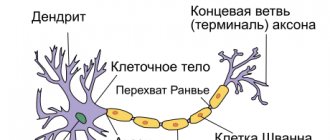
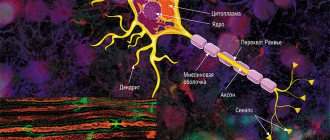
The mechanism for conducting excitation along nerve fibers depends on their type. There are two types of nerve fibers: myelinated and unmyelinated.
Metabolic processes in unmyelinated fibers do not provide rapid compensation for energy expenditure. The spread of excitation will occur with gradual attenuation - with decrement. Decremental behavior of excitation is characteristic of a low-organized nervous system. Excitation propagates due to small circular currents that arise into the fiber or into the surrounding liquid. A potential difference arises between excited and unexcited areas, which contributes to the emergence of circular currents. The current will spread from the “+” charge to the “-”. At the point where the circular current exits, the permeability of the plasma membrane for Na ions increases, resulting in depolarization of the membrane. A potential difference again arises between the newly excited area and the neighboring unexcited one, which leads to the emergence of circular currents. The excitation gradually covers neighboring areas of the axial cylinder and thus spreads to the end of the axon.
In myelin fibers, thanks to the perfection of metabolism, excitation passes without fading, without decrement. Due to the large radius of the nerve fiber due to the myelin sheath, electric current can enter and exit the fiber only in the area of interception. When stimulation is applied, depolarization occurs in the area of interception A, and the neighboring interception B is polarized at this time. Between the interceptions, a potential difference arises, and circular currents appear. Due to circular currents, other interceptions are excited, while the excitation spreads saltatory, jumpwise from one interception to another. The saltatory method of propagation of excitation is economical, and the speed of propagation of excitation is much higher (70–120 m/s) than along unmyelinated nerve fibers (0.5–2 m/s).
There are three laws for the conduction of stimulation along a nerve fiber.
Law of anatomical and physiological integrity.
Conduction of impulses along a nerve fiber is possible only if its integrity is not compromised. If the physiological properties of the nerve fiber are disrupted by cooling, the use of various drugs, compression, as well as cuts and damage to the anatomical integrity, it will be impossible to conduct a nerve impulse through it.
Law of isolated conduction of excitation.
There are a number of features of the spread of excitation in peripheral, pulpal and non-pulpate nerve fibers.
In peripheral nerve fibers, excitation is transmitted only along the nerve fiber, but is not transmitted to neighboring ones, which are located in the same nerve trunk.
In the pulpy nerve fibers, the myelin sheath plays the role of an insulator. Due to myelin, the resistivity increases and the electrical capacitance of the sheath decreases.
In non-pulp nerve fibers, excitation is transmitted in isolation. This is explained by the fact that the resistance of the fluid that fills the intercellular gaps is significantly lower than the resistance of the nerve fiber membrane. Therefore, the current that arises between the depolarized area and the unpolarized one passes through the intercellular gaps and does not enter neighboring nerve fibers.
The law of two-way conduction of excitation.
The nerve fiber conducts nerve impulses in two directions - centripetal and centrifugal.
In a living organism, excitation is carried out only in one direction. Bilateral conductivity of the nerve fiber is limited in the body by the place where the impulse originates and the valve property of synapses, which consists in the possibility of excitation in only one direction.
The composite nature of the action potential of the nerve trunk and the classification of nerve fibers
The amplitude of electrical impulses removed from the entire nerve trunk depends on the strength of the applied stimulus. A weak stimulus corresponds to a small response; as stimulation intensifies, the amplitude of the potential increases, reaching a maximum value, and then remains constant, despite a further increase in the strength of the stimulus (Fig. 47). This is explained by the fact that the electrical response of the entire nerve trunk is an algebraic sum of the action potentials of its individual fibers. In each fiber, the amplitude of the action potential does not depend on the strength of stimulation in accordance with the “all or nothing” law.
The stimulation thresholds of individual fibers differ from each other. With weak stimulus strength, excitation occurs in the most excitable superficial nerve fibers. Increasing the stimulus leads to an increase in the number of excited fibers, so the total response to stimulation increases until all fibers are involved in the reaction.
This picture can be observed if the discharge electrodes are placed on the nerve near the stimulating electrodes. As the distance between these two pairs of electrodes increases, the total action potential begins to be divided into several individual oscillations, which become most clearly expressed when the output electrodes move 10-15 cm from the site of stimulation (Fig. 48).
This phenomenon was first studied in detail by Erlanger and Gasser (1937). They showed that the reason for the division of the total action potential into components is the unequal speed of excitation along different fibers, as a result of which nerve impulses arrive at the output electrodes through these fibers non-simultaneously.
At present, nerve fibers, based on the speed of excitation, the duration of various phases of the action potential and structure, are usually divided into three main types, designated by the letters A, B and C (Fig. 49).
Type A fibers are divided into four subgroups: oc, (5, y, b. They are covered with a myelin sheath. The thickest of them are α-fibers (Aα), in warm-blooded animals and humans they have a diameter of 12-22 microns and are characterized by a significant conduction speed excitation - 70-120 m/s. Such fibers conduct excitation from the motor nerve centers of the spinal cord to the skeletal muscles (motor fibers) and from certain muscle receptors to the corresponding nerve centers. The peak action potential of the Aα fiber in warm-blooded animals lasts 0.4-0. 5 ms After its end, a trace depolarization develops, which lasts 15-20 ms and turns into a trace hyperpolarization lasting about 40-60 ms.
Three other groups of type A fibers: Ap, Aγ and Aδ - have a smaller diameter, a lower conduction velocity and a longer action potential. These are predominantly sensory fibers that conduct excitation from various receptors (tactile, some pain, temperature and receptors of internal organs) to the central nervous system.
The only exceptions are γ-fibers, a significant part of which conduct excitation in a centrifugal direction from the spinal cord cells to the so-called intrafusal muscle fibers that are part of the muscle receptors - muscle spindles (see Fig. 51, A).
Type B fibers include myelinated, predominantly preganglionic, fibers of the autonomic nervous system. The speed of excitation in these fibers in warm-blooded animals is 3-18 m/s. The duration of the action potential of type B fibers (1-2 ms) is approximately 3 times longer than the duration of the action potential of type A fibers. A distinctive feature of these fibers is that they do not exhibit a trace depolarization phase: the descending leg of the peak directly turns into a trace hyperpolarization, which in some cases it lasts more than 100 ms.
Type C fibers are non-mellose nerve fibers with a very small diameter (approximately 1 µm). The excitation speed in these fibers is no more than 3 m/s. Most type C fibers are postganglionic fibers of the sympathetic nervous system.
Type C fibers also include those nerve fibers that are involved in conducting excitation from pain receptors and some receptors of cold, heat and pressure.
Action potentials of fibers of this type are characterized by the longest duration (2 ms in warm-blooded animals). They have a long phase of trace depolarization (50-80 ms), accompanied by an even longer (300-1000 ms) trace hyperpolarization. All these data are summarized in table. 3.
Laws of conduction of excitation along nerve fibers. Classification of nerve fibers.
Laws of conduction of excitation along nerve fibers
• Law of anatomical and physiological continuity - excitation can spread along a nerve fiber only if it is morphological and functionally intact.
• The law of bilateral conduction of excitation - excitation that occurs in one part of the nerve spreads in both directions from the place of its origin. In the body, excitation always spreads along the axon from the cell body (orthodromically).
• The law of isolated conduction - excitation propagating along a fiber that is part of a nerve is not transmitted to neighboring nerve fibers.
Based on thickness and speed of nerve impulse conduction, fibers are divided into three main types: A, B and C.
Type A fibers are the thickest fibers in ns.
All type A fibers are myelinated . Their diameter varies from 3 to 22 μm (micrometers). The speed of nerve impulse conduction is from 5 to 120 m/s. The thicker the fiber, the faster the nerve impulse is carried through it. Type A fibers are divided into four types:
- Aα. Diameter from 13 to 22 microns. The speed of nerve impulse conduction is from 70 to 120 m/s. These are the thickest and fastest fibers in the nervous system. These fibers innervate skeletal muscles. These are efferent fibers of the anterior horns of the spinal cord.
- Aβ. Diameter from 8 to 13 microns. The speed of nerve impulse conduction is from 40 to 70 m/s. They belong to the afferent fibers of pressure and touch receptors.
- Aγ. Diameter from 4 to 8 microns. The speed of nerve impulse conduction is from 15 to 40 m/s. Efferent fibers of muscle spindles and part of the afferent fibers coming from pressure and touch. Also, these are the fibers of the 1st afferent neuron (? From itself).
- Aδ. Diameter from 3 to 4 microns. The speed of nerve impulse conduction is from 5 to 15 m/s. Afferent fibers from cutaneous temperature and cutaneous pain receptors. Also, partially, pressure receptors.
Type B fibers are partially myelinated . They are thinner than type A fibers. The diameter of these fibers is from 1 to 3 microns. The speed of nerve impulse conduction is from 3 to 18 m/s. These are fibers of the autonomic nervous system (lateral horns of the spinal cord) preganglionic fibers.
Snemyelized type fibers . Diameter from 0.5 to 2 microns. The speed of nerve impulse conduction is from 1.5 to 3 m/s. The fibers of the autonomic NS are postganglionic fibers and in the somatic NS there are pain and temperature fibers.
Type A fibers are more labile, i.e. the highest frequency of nerve impulse transmission. They have a very short refractory period. AP in them proceeds very quickly, approximately 0.5 ms (milliseconds).
Neuroglia, its types and significance.
Neuroglia is a large heterogeneous group of elements of nervous tissue that ensures the activity of neurons and performs nonspecific functions: supporting, trophic, delimiting, barrier, secretory and protective functions. It is an auxiliary component of nervous tissue.
In the human brain, the content of glial cells (gliocytes) is 5-10 times higher than the number of neurons, and they occupy about half of its volume. Unlike neurons, adult gliocytes are capable of division. In damaged areas of the brain, they multiply, filling defects and forming glial scars (gliosis); Tumors from glial cells (gliomas) account for 50% of intracranial neoplasms.