Классификация нервных волокон
| Амплитуда электрических импульсов, отводимых от целого нервного ствола, зависит от силы приложенного раздражения. Раздражению слабой силы соответствует небольшой потенциал действия, по мере же усиления раздражения амплитуда пика возрастает, достигает максимальной величины и затем остается постоянной, несмотря на дальнейшее увеличение силы раздражителя (рис. 156). Рис. 156. Ответы седалищного нерва лягушки на электрические стимулы возрастающей силы (по Я. Бурешу). Отведение потенциалов действия двухфазное. В начале каждой записи артефакт раздражения. Сила тока увеличивается постепенно от 1 к 6. Отметка времени 1 мсек. Объясняется это тем, что потенциал действия, отводимый от целого нервного ствола, является алгебраической суммой потенциалов отдельных его нервных волокон. В каждом волокне амплитуда потенциала действия не зависит от силы раздражения в соответствии с законом «все или ничего». Пороги же раздражения отдельных волокон отличаются друг от друга. При слабой силе стимула возбуждение возникает в наиболее возбудимых поверхностно расположенных нервных волокнах. Усиление стимула приводит к увеличению числа возбужденных волокон, поэтому суммарный пик увеличивается до тех пор, пока все волокна не вовлекаются в реакцию. Такую картину можно наблюдать, если отводящие электроды расположить на нерве вблизи раздражающих электродов. При увеличении расстояния между этими двумя парами электродов суммарный потенциал действия начинает расчленяться на несколько отдельных колебаний, которые становятся наиболее отчетливо выраженными при удалении отводящих электродов на 10—15 см от места раздражения (рис. 157). |
| Впервые это явление было подробно изучено Эрлангером и Гассером (1937). Они показали, что причиной расчленения суммарного потенциала действия на компоненты является неодинаковая скорость проведения возбуждения по разным волокнам, вследствие чего к отводящим электродам нервные импульсы поступают по этим волокнам неодновременно. Рис. 157. Комплекс составного потенциала действия смешанного нервного ствола лягушки (по Эрлангеру и Гассеру). |
| Детальный анализ показал, что между скоростью проведения импульса и диаметром нервного волокна существует примерно пропорциональная зависимость: нервные волокна проводят тем быстрее, чем они толще. В настоящее время принято подразделять нервные волокна по скорости проведения возбуждения, длительности различных фаз потенциала действия и строению на три основных типа, обозначаемых буквами А, В и С (рис. 158). Рис. 158. Схематическое изображение всех составных частей потенциала действия смешанного нерва (по Эрлангеру и Гассеру). |
Классификация нервных волокон
Нервные волокна типа А делятся на четыре подгруппы: α, β, γ и δ. Они покрыты миелиновой оболочкой. Наиболее толстые из них — так называваемые альфа-волокна (Аα) — у теплокровных животных и человека имеют диаметр 12—22 мк и характеризуются значительной скоростью проведения возбуждения: 70—120 м/сек. Такие волокна проводят возбуждение от моторных нервных центров спинного мозга к скелетным мышцам («двигательные» волокна) и от рецепторов мышц к соответствующим нервным центрам. Потенциал действия волокон Аα у теплокровных длится 0,5 мсек. После его окончания развивается следовой отрицательный потенциал, который продолжается 15—20 мсек и переходит в следовой положительный потенциал длительностью около 40—60 мсек.
Три другие группы волокон типа А: бета (Aβ)-, гамма (Аγ)- и дельта волокна имеют меньший диаметр — от 2 до 12 мк, меньшую скорость проведения и более длительный потенциал действия. Это преимущественно чувствительные волокна, проводящие возбуждение от различных рецепторов (тактильных, некоторых болевых, температурных и рецепторов внутренних органов) в центральную нервную систему. Исключение составляют лишь гамма-волокна, значительная часть которых проводит возбуждение в центробежном направлении — от клеток спинного мозга к так называемым интрафузальным мышечным волокнам которых расположены чувствительные нервные окончания — мышечные веретена.
К волокнам типа В относятся мпелиновые, преимущественно преганглионарные волокна автономной нервной системы. Скорость проведения возбуждения в этих волокнах у теплокровных животных составляет 3—14 м/сек. Продолжительность потенциала действия волокон типа В примерно в 2 раза превышает длительность потенциала действия волокон типа А. В среднем она составляет 1,2 мсек. Отличительной особенностью этих волокон является то, что в них не обнаруживается следового отрицательного потенциала: фаза реполяризации непосредственно переходит в следовой положительный потенциал, имеющий большую длительность — 100—300 мсек.
К волокнам типа С относят безмякотиые нервные волокна очень малого диаметра (порядка 0,5 мк). Скорость проведения возбуждения в этих волокнах 2—6 м/сек. Большинство С-волокон относится к постганглионарным волокнам симпатической нервной системы.
К С-волокнам относят также те нервные волокна, которые участвуют в проведении возбуждения от болевых рецепторов и некоторых рецепторов холода, тепла и давления. Потенциалы действия С-волокон характеризуются наибольшей продолжительностью (2 мсек у теплокровных животных). Они имеют длительный следовой отрицательный потенциал (50—80 мсек), сопровождающийся еще более продолжительной следовой позитивностью (300—1000 мсек).
Все эти данные суммированы в таблице.
Свойства различных нервных волокон теплокровных
| Тип волокна | Диаметр волокна (мк) | Скорость проведения (м/сек) | Длительность потенциала действия (мсек) | Длительность отрицательного следового потенциала (мсек) | Длительность положительного следового потенциала (мсек) | Функция |
| А (α) | 12-22 | 70-120 | 0,4-0,5 | 12-20 | 40-60 | Моторные волокна скелетных мышц, афферентные волокна мышечных рецепторов |
| А (β) | 8-12 | 40-70 | 0,4-0,6 | — | — | Афферентные волокна от рецепторов прикосновения |
| А (γ) | 4-8 | 15-40 | 0,5-0,7 | — | — | Афферентные волокна от рецепторов прикосновения и давления, эфферентные волокна к мышечным веретенам |
| А (Δ) | 1-4 | 5-15 | 0,6-1,0 | — | — | Афферентные волокна от некоторых рецепторов тепла, давления, боли |
| В | 1-3 | 3-14 | 1-2 | Отсут- ствует | 100-300 | Преганглионарные вегетативные волокна |
| С | 0,5-1,0 | 0,5-2 | 2,0 | 50-80 | 300-1000 | Преганглионарные вегетативные волокна, афеерентные волокна от некоторых рецепторов боли, давления, тепла |
Нормальная физиология: конспект лекций
2. Механизмы проведения возбуждения по нервному волокну. Законы проведения возбуждения по нервному волокну
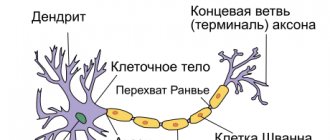
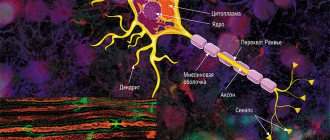
Механизм проведения возбуждения по нервным волокнам зависит от их типа. Существуют два типа нервных волокон: миелиновые и безмиелиновые.
Процессы метаболизма в безмиелиновых волокнах не обеспечивают быструю компенсацию расхода энергии. Распространение возбуждения будет идти с постепенным затуханием — с декрементом. Декрементное поведение возбуждения характерно для низкоорганизованной нервной системы. Возбуждение распространяется за счет малых круговых токов, которые возникают внутрь волокна или в окружающую его жидкость. Между возбужденными и невозбужденными участками возникает разность потенциалов, которая способствует возникновению круговых токов. Ток будет распространяться от «+» заряда к «—». В месте выхода кругового тока повышается проницаемость плазматической мембраны для ионов Na, в результате чего происходит деполяризация мембраны. Между вновь возбужденным участком и соседним невозбужденным вновь возникает разность потенциалов, что приводит к возникновению круговых токов. Возбуждение постепенно охватывает соседние участки осевого цилиндра и так распространяется до конца аксона.
В миелиновых волокнах благодаря совершенству метаболизма возбуждение проходит, не затухая, без декремента. За счет большого радиуса нервного волокна, обусловленного миелиновой оболочкой, электрический ток может входить и выходить из волокна только в области перехвата. При нанесения раздражения возникает деполяризация в области перехвата А, соседний перехват В в это время поляризован. Между перехватами возникает разность потенциалов, и появляются круговые токи. За счет круговых токов возбуждаются другие перехваты, при этом возбуждение распространяется сальтаторно, скачкообразно от одного перехвата к другому. Сальтаторный способ распространения возбуждения экономичен, и скорость распространения возбуждения гораздо выше (70—120 м/с), чем по безмиелиновым нервным волокнам (0,5–2 м/с).
Существует три закона проведения раздражения по нервному волокну.
Закон анатомо-физиологической целостности.
Проведение импульсов по нервному волокну возможно лишь в том случае, если не нарушена его целостность. При нарушении физиологических свойств нервного волокна путем охлаждения, применения различных наркотических средств, сдавливания, а также порезами и повреждениями анатомической целостности проведение нервного импульса по нему будет невозможно.
Закон изолированного проведения возбуждения.
Существует ряд особенностей распространения возбуждения в периферических, мякотных и безмякотных нервных волокнах.
В периферических нервных волокнах возбуждение передается только вдоль нервного волокна, но не передается на соседние, которые находятся в одном и том же нервном стволе.
В мякотных нервных волокнах роль изолятора выполняет миелиновая оболочка. За счет миелина увеличивается удельное сопротивление и происходит уменьшение электрической емкости оболочки.
В безмякотных нервных волокнах возбуждение передается изолированно. Это объясняется тем, что сопротивление жидкости, которая заполняет межклеточные щели, значительно ниже сопротивления мембраны нервных волокон. Поэтому ток, возникающий между деполяризованным участком и неполяризованным, проходит по межклеточным щелям и не заходит при этом в соседние нервные волокна.
Закон двустороннего проведения возбуждения.
Нервное волокно проводит нервные импульсы в двух направлениях — центростремительно и центробежно.
В живом организме возбуждение проводится только в одном направлении. Двусторонняя проводимость нервного волокна ограничена в организме местом возникновения импульса и клапанным свойством синапсов, которое заключается в возможности проведения возбуждения только в одном направлении.
Составной характер потенциала действия нервного ствола и классификации нервных волокон
Амплитуда электрических импульсов, отводимых от целого нервного ствола, зависит от силы приложенного раздражителя. Раздражителю слабой силы соответствует небольшой ответ; по мере усиления раздражения амплитуда потенциала возрастает, достигая максимальной величины, и затем остается постоянной, несмотря на дальнейшее увеличение силы раздражителя (рис. 47). Объясняется это тем, что электрический ответ целого нервного ствола является алгебраической суммой потенциалов действия отдельных его волокон. В каждом волокне амплитуда потенциала действия не зависит oт силы раздражения в соответствии с законом «все или ничего».
Пороги раздражения отдельных волокон отличаются друг от друга. При слабой силе стимула возбуждение возникает в наиболее возбудимых поверхностно расположенных нервных волокнах. Усиление стимула приводит к увеличению числа возбужденных волокон, поэтому суммарный ответ на раздражение увеличивается до тех пор, пока все волокна не вовлекаются в реакцию.
Такую картину можно наблюдать, если отводящие электроды расположить на нерве вблизи раздражающих электродов. При увеличении расстояния между этими двумя парами электродов суммарный потенциал действия начинает расчленяться на несколько отдельных колебаний, которые становятся наиболее отчетливо выраженными при удалении отводящих электродов на 10—15 см от места раздражения (рис. 48).
Впервые это явление было подробно изучено Эрлангером и Гассером (1937). Они показали, что причиной расчленения суммарного потенциала действия на компоненты является неодинаковая скорость проведения возбуждения по разным волокнам, вследствие чего к отводящим электродам нервные импульсы поступают по этим волокнам неодновременно.
В настоящее время нервные, волокна по скорости проведения возбуждения, длительности различных фаз потенциала действия и строению принято подразделять на три основных типа, обозначаемых буквами А, В и С (рис. 49).
Волокна типа А делятся на четыре подгруппы: ос, (5, у, б. Они покрыты миелиновой оболочкой. Наиболее толстые их них α-волокна (Аα), у теплокровных животных и человека они имеют диаметр 12—22 мкм и характеризуются значительной скоростью проведения возбуждения — 70—120 м/с. Такие волокна проводят возбуждение от моторных нервных центров спинного мозга к скелетным мышцам (двигательные волокна) и от определенных рецепторов мышц к соответствующим нервным центрам. Пик потенциала действия волокна Аα у теплокровных длится 0,4—0,5 мс. После его окончания развивается следовая деполяризация, которая продолжается 15—20 мс и переходит в следовую гиперполяризацию длительностью около 40—60 мс.
Три другие группы волокон типа А: Ар, Аγ и Аδ — имеют меньший диаметр, меньшую скорость проведения и более длительный потенциал действия. Это преимущественно чувствительные волокна, проводящие возбуждение от различных рецепторов (тактильных, некоторых болевых, температурных и рецепторов внутренних органов) в ЦНС.
Исключение составляют лишь γ-волокна, значительная часть которых проводит возбуждение в центробежном направлении от клеток спинного мозга к так называемым интрафузальным мышечным волокнам, входящим в состав рецепторов мышц — мышечных веретен (см. рис. 51, А).
К волокнам типа В относятся миелинизированные, преимущественно преганглионарные, волокна вегетативной нервной системы. Скорость проведения возбуждения в этих волокнах у теплокровных животных составляет 3—18 м/с. Продолжительность потенциала действия волокон типа В (1—2 мс) примерно в 3 раза превышает длительность потенциала действия волокон типа А. Отличительной особенностью этих волокон является то, что в них не обнаруживается фаза следовой деполяризации: нисходящее колено пика непосредственно переходит в следовую гиперполяризацию, которая в ряде случаев продолжается более 100 мс.
К волокнам типа С относят безмякотные нервные волокна очень малого диаметра (примерно 1 мкм). Скорость проведения возбуждения в этих волокнах не более 3 м/с. Большинство волокон типа С — это постганглионарные волокна симпатической нервной системы.
К волокнам типа С относят также те нервные волокна, которые участвуют в проведении возбуждения от болевых рецепторов и некоторых рецепторов холода, тепла и давления.
Потенциалы действия волокон этого типа характеризуются наибольшей продолжительностью (2 мс у теплокровных животных)., Они имеют длительную фазу следовой деполяризации (50—80 мс), сопровождающуюся еще более продолжительной (300 — 1000 мс) следовой гиперполяризацией. Все эти данные суммированы в табл. 3.
Законы проведения возбуждения по нервным волокнам. Классификация нервных волокон.
Законы проведения возбуждения по нервным волокнам
• Закон анатомической и физиологической непрерывности – возбуждение может распространяться по нервному волокну только в случае его морфологической и функциональной целостности.
• Закон двустороннего проведения возбуждения – возбуждение, возникающее в одном участке нерва, распространяется в обе стороны от места своего возникновения. В организме возбуждение всегда распространяется по аксону от тела клетки (ортодромно).
• Закон изолированного проведения – возбуждение, распространяющееся по волокну, входящему в состав нерва, не передается на соседние нервные волокна.
По толщине и по скорости проведения нервного импульса волокна делятся на три основных типа: A, B и С.
Волокна типа А самые толстые волокна в нс.
Все волокна типа А миелинизированные. Их диаметр варьируется от 3 до 22 мкм (микрометров). Скорость проведения нервного импульса – от 5 до 120 м/с.Чем толще волокно, тем быстрее проведение по нему нервного импульса. Волокна типа А делятся на четыре вида:
- Аα. Диаметр от 13 до 22 мкм. Скорость проведения нервного импульса от 70 до 120 м/с. Это самые толстые и быстрые волокна в нервной системы. Эти волокна иннервируют скелетные мышцы. Это эфферентные волокна передних рогов спинного мозга.
- Аβ. Диаметр от 8 до 13 мкм. Скорость проведения нервного импульса от 40 до 70 м/с. Они относятся к афферентным волокнам рецепторов давления и прикосновения.
- Аγ. Диаметр от 4 до 8 мкм. Скорость проведения нервного импульса от 15 до 40 м/с. Эфферентные волокна мышечных веретен и часть афферентных волокон идущих от давления и прикосновения. Также, это волокна 1-го афферентного нейрона (? От себя).
- Аδ. Диаметр от 3 до 4 мкм. Скорость проведения нервного импульса от 5 до 15 м/с. Афферентные волокна от кожных температурных и кожных болевых рецепторов. Также, частично, рецепторы давления.
Волокна типа В частично миелинизированные. Они более тонкие, чем волокна типа А. Диаметр этих волокон от 1 до 3 мкм. Скорость проведения нервного импульса от 3 до 18 м/с. Это волокна вегетативной нервной системы (боковые рога спинного мозга) преганглионарные волокна.
Волокна типа Снемиелизированные. Диметр от 0.5 до 2 мкм. Скорость проведения нервного импульса от 1.5 до 3 м/с. Волокна вегетативной НС постганглионарные волокна и в соматической НС болевые и температурные.
Волокна типа А более лабильны, т.е. наиболее высокая частота передачи нервного импульса. У них очень короткое время рефрактерного периода. ПД в них протекает очень быстро, примерно 0.5 мс (миллисекунды).
Нейроглия, ее виды и значение.
Нейроглия — обширная гетерогенная группа элементов нервной ткани, обеспечивающая деятельность нейронов и выполняющая неспецифические функции: опорную, трофическую, разграничительную, барьерную, секреторную и защитную функции. Является вспомагательнымкомпанентом нервной ткани.
В мозге человека содержание глиальных клеток (глиоцитов) в 5-10 раз превышает число нейронов, причем они занимают около половины его объема. В отличие от нейронов, глиоциты взрослого способны к делению. В поврежденных участках мозга они размножаются, заполняя дефекты и образуя глиальные рубцы (глиоз); опухоли из клеток глии (глиомы) составляют 50% внутричерепных новообразований.