Structure and functions of a neuron[edit | edit code]
A. Structure and functions of a nerve cell
Excitable cells respond to stimuli by changing the state of their membranes. There are two types of excitable cells: nerve cells, which conduct and transduce impulses in the nervous system, and muscle cells, which contract either in response to nerve impulses or autonomously.
Nervous system
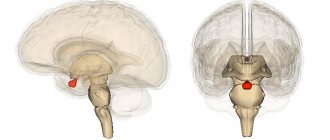
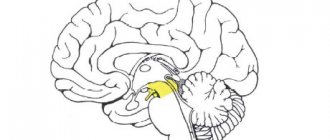
The human body consists of more than 1010 nerve cells, or neurons.
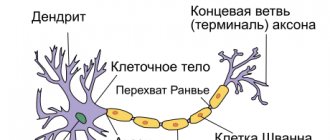
A neuron
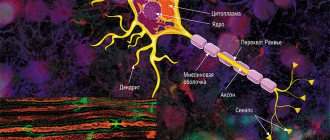
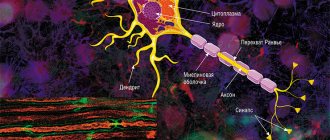
is a structural and functional unit of the nervous system. A typical neuron (motoneuron, A1) consists of a soma, or cell body, and two types of processes - axon and dendrites. In addition to the usual cellular organelles, such as the nucleus and mitochondria (A2), the neuron contains neurofibrils and neurotubules. A neuron receives afferent signals (excitatory and inhibitory) from several and sometimes several thousand neighboring neurons through dendrites (usually tree-like), and the signals are summed along the neuron body on the cell membrane (summation).The axon starts from the axon hillock of the neuron body: it carries out transmission of efferent nerve signals to nearby or distant effectors (muscle and secretory cells) and nearby neurons. Axons often have branches (collaterals), which branch further and end in swellings - synaptic vesicles or synaptic endings. If the total potential at the axon hillock exceeds a certain threshold, an action potential is generated and transmitted down the axon, where it reaches the next synapse through the synaptic terminal (A1, 3), described below.
Vesicles containing various substances (proteins, lipids, sugars and neurotransmitter molecules) are transported from the Golgi complex in the soma to the synaptic terminal and to the tips of dendrites by rapid axonal transport (40 cm/day). This type of anterograde (forward-directed) transport along neurotubules is carried out by kinesin (myosin-like protein), and the energy required for this is supplied by ATP. Endogenous and exogenous substances, such as nerve growth factor (NGF), herpes virus, polio virus, and tetanus toxin, are transported retrogradely from peripheral sites to the soma at a rate of ~25 cm/day. Slow axonal transport (~1 mm/day) plays an important role in the treatment of severe neuritis.
The plasma membrane of the soma continues along the axon and is called the axolemma (A1, 2).
In the central nervous system (CNS), the axolemma is surrounded by oligodendrocytes, and in the peripheral nervous system, by Schwann cells (A1, 2). A nerve fiber consists of an axon and its sheath. In some neurons, Schwann cells form a multilayered myelin sheath of double phospholipid layers (A1, 2) around the axon, which insulates the axon from ionic currents. The myelin sheath is interrupted approximately every 1.5 mm at the nodes of Ranvier (A1). The conductivity of myelinated nerve fibers is much higher than that of unmyelinated nerve fibers and increases with the diameter of the nerve fiber.
A synapse (A3) is the site where a neuron's axon interacts with effectors or other neurons. Synaptic transmission in almost all mammals is carried out by chemical compounds rather than by electrical signals. In response to an electrical signal in the axon, neurotransmitters are released from vesicles on the presynaptic membrane by exocytosis. The transmitter diffuses through the synaptic cleft (10-40 nm) to the postsynaptic membrane, where it connects with receptors that create new electrical signals (AZ). Depending on the type of neurotransmitter and receptor involved in the process, the neurotransmitter has either an excitatory (for example, acetylcholine in skeletal muscle) or an inhibitory effect (for example, glycine in the central nervous system) on the postsynaptic membrane. Because the postsynaptic membrane does not normally release neurotransmitters (there are only a few exceptions), nerve impulses can only pass through the synapse in one direction. Thus, the synapse acts as a valve that ensures orderly signal transmission. Synapses are also sites at which the transmission of a nerve impulse can be converted by other (excitatory or inhibitory) neurons.
To enter into the problem of neuron activity in the CNS (central nervous system), it is necessary to briefly dwell on its morphology. Neuron membrane. Neurons have a high level of morphological and functional specialization, and the first level of specialization is manifested in the structure and dynamics of its membrane.
As the results of electron microscopy show, the plasma membrane of neurons has the same structure as that of somatic cells: it is built from lipids and proteins (chains of amino acids).
The main structure of the membrane is two-layer and is a “sandwich” of phospholipids, which are arranged in such a way that the polar (charged) parts are adjacent to the outer part of the membrane, and the uncharged parts are directed into the cell.
This organization maximizes the number of hydrophobic and hydrophilic compounds that can form and make a relatively strong and very thin shell impermeable to most polar molecules or ions.
The membrane is dynamic and is often described as a “fluid mosaic” structure (Singer and Nicolson, 1972).
Lipids diffuse freely from one area to another, thereby providing the membrane with the properties of a liquid.
Two liquid lipid layers of the membrane (outer and inner) allow specialized proteins to float freely in it and perform their functions.
Proteins can penetrate both layers, forming channels for the transport of ions and small molecules through them.
Such “integral proteins” often form intermembrane structures.
Others, “peripheral proteins,” are localized only in the outer or inner membranes; they are mobile and perform specific functions.
In a liquid membrane, proteins are often thought of as membrane particles floating randomly in a sea of lipids.
It should be noted that proteins that penetrate both layers of the membrane, being one end outside and the other inside the cell, transform sections of the membrane into functional units that provide the specific needs of the neuron.
Diagram of the structure of a neuron membrane
The neuron membrane is heterogeneous; some authors identify specific areas in it, such as the dendritic zone with a large number of synaptic contacts and the presynaptic zone of the axon. In addition, the neuron membrane is asymmetrical.
On its outer part (as well as on the membrane of somatic cells) there is a large amount of sialic acid carbohydrates, which provide a negative charge on the outer surface of the membrane.
Membrane proteins.
According to their function, they are divided into pumps, channels, receptors, enzymes and structural proteins.
Pumps move ions and molecules against concentration gradients and maintain their required concentrations in the cell.
Since charged molecules cannot pass through the lipid bilayer, the membranes have a set of specific protein channels through which certain ions pass into the cell.
Cell membranes, with the help of receptor proteins, recognize and attach different molecules to themselves.
Enzymes are located in or on the membrane and facilitate chemical reactions at the surface of the membrane, such as ATPase, which breaks down ATP, the universal unit of fuel, to provide heat for local chemical processes.
Structural proteins ensure the connection of cells into organs and the maintenance of subcellular structure.
Not all membrane proteins have a strictly fixed function.
Some proteins can perform simultaneously the functions of a receptor, an enzyme, and a pump.
In addition to ion pumps and tubules, neurons also require other proteins to perform basic functions.
One of these proteins is the enzyme adenylate cyclase, which regulates the intracellular concentration of cyclic adenosine monophosphate (cyclic AMP - cAMP).
Cyclic nucleotides, such as cAMP, are called “second messengers.” Inside the cell, cAMP “collects” information from primary messengers (neurotransmitters) and prepares the cytoplasm for possible changes in its metabolism.
The main hypothesis is that an increase in cAMP concentration in response to the arrival of neurotransmitters such as norepinephrine and dopamine at postsynaptic receptors leads to an increase in protein kinase activity, which (1) phosphorylates certain membrane proteins and changes membrane permeability, (2) alters metabolic processes in the cell due to the activation and induction of certain enzymes and proteins.
General conclusion: cAMP somehow sets the level of excitability of the neuron (Cotman, McGaugh, 1980)
Understanding the functions of membrane proteins is one of the steps towards understanding the functions of a neuron.
Like all other cells of the body, the neuron maintains a constant internal environment, which differs significantly from the intercellular fluid surrounding the neuron.
The differences in the concentrations of sodium and potassium ions (Na*, Ka*) are especially pronounced. The external environment is approximately 10 times richer in Na* than the internal environment, and the internal environment is 10 times richer in Ka* than the intercellular fluid. This difference in the concentration of sodium and potassium ions underlies the maintenance and development of electrical potentials on the membrane of nerve cells.
2. Core. Each nerve cell has a nucleus that stores genetic material in the form of chromosomes. Chromosomes are made up of deoxyribonucleic acid (DNA) and proteins, which together form genes.
During embryonic development, genes control protein synthesis and, through proteins, ensure cell differentiation, its final shape and synaptic connections with other cells.
In the mature state of the neuron, genes, through the control of protein synthesis, control the activity of the neuron.
The nucleus is separated from the cytoplasm by two membranes, which converge in some places and form pores through which the exchange of substances takes place between the cytoplasm and the contents of the nucleus. 3. Mitochondria. A neuron needs a lot of energy to perform its functions.
The macroergic molecule ATP (adenosine triphosphoric acid) is the main source of energy.
Motile and plastic mitochondria are organelles surrounded by a double membrane in which ATP synthesis occurs.
Typically, mitochondria are small (0.5-3 microns long) intracellular formations, which, due to their mobility, are located in those places where energy is needed to maintain chemical processes.
To provide intracellular chemical processes with heat, ATP is split (hydrolyzed): the phosphorus ion and the energy that held this ion with the ADP (adenosine diphosphoric acid) ion are separated from it.
Energy is used to maintain chemical processes in the neuron, and ADP-3 and P*3 enter the mitochondria, in which the energy released from the oxidation of glucose is used to combine these ions and form ATP.
Internal contents of the cell body: R - nucleus, P - polysomes, T - microtubules, M - mitochondria, MF - microfilaments, R-ER - rough endoplasmic reticulum.
4. Construction proteins-ribosomes, rough endoplasmic reticulum and Golgi apparatus
These structures ensure protein synthesis in the neuron and its integration into membrane structures.
The proteins of neurons, like other cells, need constant renewal.
Without protein renewal, a neuron can survive for several days.
The cytoplasm of the neuron body contains a large number of ribosome clusters.
Ribosomes are about 4 nm in diameter and are formed from proteins and ribonucleic acid.
Clusters of ribosomes, called polysomes, carry out the synthesis of soluble proteins, including enzymes, in the cytoplasm.
Individual ribosomes in polysomes are associated with messenger RNA (mRNA). mRNA is a long chain of nucleic acids represented by four nucleotides: adenine, guanine, cytosine and uracil.
The sequence of this nucleotide chain encodes the sequence of amino acids in the synthesized protein.
A special transfer RNA (tRNA) “recognizes” a specific trinucleotide chain on messenger RNA (mRNA) and binds a specific amino acid to it.
As protein is synthesized, the mRNA moves through the ribosome and different amino acids are sequentially attached to its trinucleotide tRNA chains until the mRNA runs out.
Then the chain of amino acids enters the cytoplasm (Lehninger, 1982)
The synthesis of membrane proteins and their incorporation into membranes is carried out with the help of the rough surface endoplasmic reticulum (rough ER), smooth endoplasmic reticulum (smooth ER) and the Golgi apparatus.
The rough ER is a labyrinthine system of membrane tubes, vesicles and cisterns, whose surface protruding into the interior of the neuron is strewn with ribosomes linked to each other using mRNA.
Hence the term “rough surface”.
Membrane proteins insert into the rough ER.
The smooth ER is a continuation of the rough ER and lacks ribosomes.
The smooth ER is involved in protein distribution throughout the neuron: namely, it transports newly synthesized proteins to the dendrites.
From the smooth ER, proteins are transported to the Golgi apparatus, where they can be modified, for example, if this protein is a glycoprotein, then a carbohydrate is added to it.
Proteins in the Golgi apparatus are concentrated and then “packaged” in membrane vesicles and sequestered for subsequent delivery to other parts of the cell.
Next to the Golgi apparatus there are “clouds” of small vesicles, which possibly transport synthesized proteins to different parts of the neuron. Lysosomes. Lysosomes belong to the intracellular digestive system. This structure, like the reticulum, is enclosed in a membrane.
Lysosomes do not have a specific shape or size.
They contain a variety of hydrolytic enzymes that break down and digest a variety of compounds found both inside and outside the cell.
Digested substances can be intracellular, and such digestion is called autophagy. Digestion of extracellular substances by lysosomes is called heterophagy. Cytoskeletal network.
The body and processes of the neuron contain an extensive cytoskeletal network consisting of microtubules, neurofilaments and microfilaments.
They pass through the entire neuron, connecting all its parts.
This network is the framework of the neuron, maintaining its specific shape.
On the other hand, the cytoskeletal network performs a transport function.
In the cell body, microtubules and smaller tubes, neurofilaments and microfilaments, occupy most of it, not occupied by other organelles, and from the neuron body, all these tubes penetrate into the dendrites and axons.
Microtubules consist of long, unbranched tubes of varying lengths.
Their walls are built from subunits of a specific protein - tubulin (from the Latin tubula - tube).
Neurofilaments are thinner than microtubules.
They also have a tubular structure and are found only in neurons.
It has been shown that in large axons there are significantly more of them than microtubules, while in small axons and dendrites their ratio is opposite.
Neurofilaments and their relationship with microtubules change with aging.
In Alzheimer's disease they turn into tangles and plaques.
It is assumed that microtubules and neurofilaments in axons and dendrites perform a transport function between the body and processes of the neuron in both directions: from the body to the processes - anterograde transport, from the processes to the body - retrograde transport.
This hypothesis was confirmed experimentally.
After injection of labeled amino acids near the cell body, it was shown by radiography that these amino acids are taken up by the cell bodies of neurons and incorporated into protein, which is then transported along the axon and its collaterals.
In these experiments, two types of axonal transport were identified: slow transport, proceeding at a speed of 1 mm per day, and fast, proceeding at a speed of several hundred millimeters per day.
Many transported substances are associated with synaptic transmission functions.
Microfilaments are present in large quantities in nerve processes.
They are abundant in neuroglia and are involved in some connections between neurons (Shepherd, 1997).
Dendrites.
Dendrites, with all their diversity among neurons, contain the same organelles, except for the nucleus, as the body.
In most dendrites, a large number of microtubules run in parallel.
On the other hand, dendrites contain only a small amount of neurofilaments.
Mitochondria are oriented along the dendrite and reach a length of 9 µm.
The rough ER is well expressed only at the beginning of the dendrite; as it moves away from the body, the ER disappears.
Smooth ER is distributed along the entire length of the dendrites.
It runs parallel to microtubules and microfilaments, forming small protrusions along its course.
The smooth ER is believed to be distributed throughout the dendrite and transport various chemicals.
The bulk of the protein is synthesized in the cell body, but some proteins are synthesized in the proximal regions of the dendrites, where a large mass of smooth ER is localized.
These proteins are transported to the distal dendrites by cisternae and smooth ER along the surface of the microtubule system.
Proteins that are synthesized in the cell body are also transported to dendrites using smooth ER.
The dendrites contain a large number of spines, on which there are mainly axodendritic synapses.
For example, on the dendrites of pyramidal neurons, on average, about 4000 spines are localized, which is approximately 43% of the total synaptic contacts of these neurons (P (Cotman, McGaugh, 1980).
Each spine is a protrusion on a dendrite about 2 μm long, which consists of a thin neck ending in an ovoid protrusion.
The cytoplasm of the spines is filled with thin filaments and a very small number of microtubules. As mentioned above, spines are included in synaptic structures.
Axon. Unlike dendrites, an axon is usually a single process.
It does not have rough ER or ribosomes, but it contains mitochondria, a large number of neurofilaments, microtubules, smooth ER and a small number of lysosomes.
Part of the smooth ER, cisterns of different shapes, are connected to each other by thin tubes, and this system runs along the entire axon.
The interior of the axon is filled with jelly-like axoplasm, which supports its cytoskeletal network.
In the vast majority of large neurons, the axons are covered throughout their entire length with a sheath called myelin.
Myelin never covers dendrites.
It is known that myelin is produced from the plasma membrane of glial cells that wrap around the axon.
In the peripheral nervous system, axons wrap around special glial, or Schwann, cells.
The exposed areas of the axon between Schwann cells are called nodes of Ranvier.
In the central nervous system, myelin sheaths are formed by oligodendrocytes.
Action potentials travel quickly along myelinated axons due to saltatory (jumping) movement from one node of Ranvier to another. Synapse. Neuromuscular junction.
All synapses have the same structure.
The presynaptic and postsynaptic membranes are highly specialized at the site of contact and form the synaptic connection.
The presynaptic plaque, or axon terminal, contains vesicles with a diameter of about 20-65 nm, which are filled with a substance - a neurotransmitter.
The presence of vesicles and synaptic connection is an indicator of a chemical synapse.
The general details of the structure are the same for all synapses, but the fine structure of the synapses depends on the characteristics of the pre- and postsynaptic neurons; this structure is significantly different in the neuromuscular and interneuron synapses.
Neuromuscular junction: A - axon, B - synaptic endings of the axon in the form of plaques, M - muscle, mt - mitochondria, sp - synaptic vesicles, a - active zone, Sh - synaptic cleft, ss - connecting folds of the muscle cell membrane.
Immediately before the neuromuscular junction, the myelin sheath on the axon disappears, and then the axon, surrounded only by a thin layer of Schwann cell cytoplasm, divides into branches (collaterals).
At the site of synaptic contact between the axon collateral and the muscle cell, the membrane of the latter forms many folds - connecting folds.
The space between the presynaptic and postynaptic parts of the synapse is called the synaptic cleft. The synaptic cleft at the neuromuscular junction is wider than between neurons in the central nervous system.
The synaptic terminals of axons contain several mitochondria, a network of fibrous protein, and many synaptic vesicles containing a neurotransmitter, namely up to 10,000 molecules of acetylcholine, which causes an excitatory effect on the muscle cell membrane.
Some vesicles are distributed randomly along the axon branch, while others are located along the presynaptic region - the active zone, from which they are released into the synaptic cleft.
Inside the two-layer plasma membrane in the active zone there are intramembrane particles with areas to which vesicles are attached.
As the action potential travels along the axon, the vesicles float to these areas and merge with them.
Opposite the active zone of the presynaptic membrane are the functional folds of the muscle cell membrane.
Inside the membrane, mainly in the crests of the folds, there is a large number of intramembrane inclusions, with which the neurotransmitter apparently connects. Synapses of the central nervous system. Each synapse in the central nervous system is formed by two neurons - pre- and postsynaptic.
As in the neuromuscular junction, these synapses have pre- and postsynaptic membranes, between which there is a synaptic cleft about 20-50 nm wide.
The presynapse contains neurotransmitter vesicles, mitochondria, membrane cisterns, random microtubules, and many fibers.
There are two types of synapses in the central nervous system.
CNS synapses differ from neuromuscular junction synapses in several respects.
If in the neuromuscular junction acetylcholine has an excitatory effect on the muscle cell membrane, then in the central nervous system in some interneuron synapses acetylcholine has an excitatory effect on the neuron membrane, and in other synapses it has an inhibitory effect.
While at neuromuscular junctions there is only one neurotransmitter, at the synapses of the central nervous system there are more than 100 different neurotransmitters.
The first type of synapse has a fairly typical shape.
The presynaptic membrane of the axon has many compacted projections onto its cytoplasmic part, and the postsynaptic membrane of the dendrite also has compactions, but continuous ones; on the cytoplasmic surface there is a postsynaptic compaction. The typical length of such a synaptic contact is approximately 5 microns. Type 1 synapses are the most numerous; they are always axodendritic, formed on dendrite spines and never formed on the neuron body.
The condensed projections of the presynaptic membrane are a series of pyramids organized into hexagonal structures. The active zones of synapses are located in the spaces between these structures. The active zones contain synaptic vesicles, as well as many intramembrane inclusions.
Postsynaptic condensations are fibrous tissue that resembles a roughly woven carpet.
This tissue lies opposite the postsynaptic membrane.
In these seals, thin fibers grow together, which are touched by microtubules containing small inclusions.
The outer surface of the postsynaptic membrane overlying this seal is sensitive to a specific neurotransmitter; many bristles and fibers are oriented in the direction of the synaptic cleft, and some of them connect to the presynaptic membrane.
The second type of synapse is never formed on spines.
They are usually localized on the cell bodies of neurons.
It is assumed that, unlike type 1 synapses, type 2 synapses are inhibitory synapses. The main feature of type 2 synapses is the absence of protruding postsynaptic density; The plasma membrane region at the synapse is very thin.
Artificial stimulation of a nerve cell[edit | edit code]
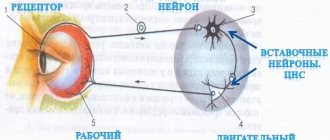
When an electrical impulse from an external source is applied to a nerve cell, current flows from the positively charged electrode (anode) and exits to the negatively charged electrode [cathode]. The nerve fiber below the cathode depolarizes and, provided the threshold potential is reached, an action potential is generated.
The speed of conduction of an impulse along a nerve can be measured by placing two electrodes on the skin along the nerve at a known distance from each other, then stimulating that nerve (containing numerous neurons) and recording the time it takes for the total action potential to travel the distance between the electrodes. The speed of signal transmission in humans is usually from 40 to 70 m/s. Values below 40 m/s are considered pathological.
- Accidental exposure to electricity. High voltage, especially low-frequency alternating current (for example, from an electrical outlet), and also under conditions of reduced resistance (bare feet, accident in the bathroom), mainly affects the conduction of signals in the heart, which can cause ventricular fibrillation.
Direct current usually acts as a stimulus only when switched on and off: high-frequency alternating current (> 15 kHz), in contrast, is not capable of causing depolarization, but damages body tissues. Diathermy is based on this principle.