Строение и функции нейрона[править | править код]
А. Строение и функции нервной клетки
Возбудимые клетки реагируют на раздражители путем изменения состояния мембран. Существуют два типа возбудимых клеток: нервные клетки, которые проводят и преобразуют импульсы в нервной системе, и мышечные клетки, которые сокращаются либо в ответ на нервные импульсы, либо автономно.
Нервная система
человека состоит из более чем 1010 нервных клеток, или нейронов.
Нейрон
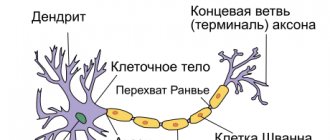
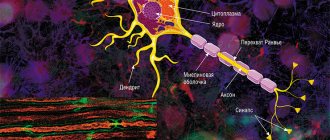
— это структурная и функциональная единица нервной системы. Типичный нейрон (мотонейрон, А1) состоит из сомы, или тела клетки, и двух типов отростков -аксона и дендритов. Кроме обычных клеточных органелл, таких как ядро и митохондрии (А2), в нейроне есть нейрофибриллы и нейротрубочки. Нейрон получает афферентные сигналы (возбуждающие и тормозящие) от нескольких, а иногда и от нескольких тысяч соседних нейронов через дендриты (обычно древовидные], и сигналы суммируются вдоль тела нейрона на клеточной мембране (суммация). Аксон начинается от аксонного холмика тела нейрона: он осуществляет передачу эфферентных нервных сигналов к ближайшим либо отдаленным эффекторам (мышечным и секреторным клеткам) и близлежащим нейронам. Аксоны часто имеют ответвления (коллатерали), которые ветвятся далее и заканчиваются вздутиями — синаптическими пузырьками или синаптическими окончаниями. Если суммарный потенциал у аксонного холмика превышает некоторый порог, генерируется потенциал действия, который передается по аксону вниз, где достигает следующего синапса через синаптическое окончание (А1, 3), описанное ниже.
Везикулы, содержащие различные вещества (белки, липиды, сахара и молекулы медиаторов), транспортируются от комплекса Гольджи в соме к синаптическому окончанию и к кончикам дендритов путем быстрого аксонного транспорта (40 см/сутки). Этот вид антероградного (направленного вперед) транспорта по ходу нейротрубочек осуществляется кинезином (миозино-подобным белком), а энергия, необходимая для этого, поставляется АТФ. Эндогенные и экзогенные вещества, такие как фактор роста нервов (ФРН, или NRF), вирус герпеса, вирус полиомиелита и столбнячный токсин, проводятся ретроградным (направленным назад) транспортом от периферических участков к соме со скоростью ~ 25 см/сутки. Медленный аксонный транспорт (~ 1 мм/сутки) играет важную роль при лечении тяжелых невритов.
Плазматическая мембрана сомы продолжается вдоль аксона и называется аксолеммой (А1, 2).
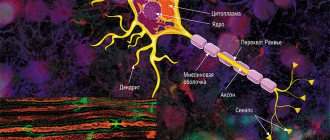
В центральной нервной системе (ЦНС) аксолемма окружена олигодендроцитами, а в периферической — шванновскими клетками (А1, 2). Нервное волокно состоит из аксона и его оболочки. В некоторых нейронах шванновские клетки образуют вокруг аксона многослойную миелиновую оболочку из двойных фосфолипидных слоев (А1, 2), которая изолирует аксон от ионных токов. Миелиновая оболочка прерывается примерно каждые 1,5 мм у перехватов Ранвье (А1). Проводимость миелинизированных нервных волокон гораздо выше, чем немиелинизированных, и увеличивается вместе с диаметром нервного волокна.
Синапс (А3) — это участок, где аксон нейрона взаимодействует с эффекторами или другими нейронами. Синаптическая передача почти у всех млекопитающих осуществляется с помощью химических соединений, а не с помощью электрических сигналов. В ответ на электрический сигнал в аксоне из везикул на пресинаптической мембране происходит высвобождение нейромедиаторов путем экзоцитоза. Медиатор диффундирует через синаптическую щель (10-40 нм) к постсинаптической мембране, где он соединяется с рецепторами, создающими новые электрические сигналы (АЗ). В зависимости от типа участвующих в процессе нейромедиатора и рецептора нейромедиатор оказывает на постсинаптическую мембрану или возбуждающий (например, ацетилхолин в скелетной мышце), или тормозящий эффект (например, глицин в ЦНС). Поскольку постсинаптическая мембрана в норме не высвобождает нейромедиаторы (существует всего несколько исключений), нервные импульсы могут пройти через синапс только в одном направлении. Таким образом, синапс действует как клапан, который обеспечивает упорядоченную передачу сигнала. Синапсы являются также участками, в которых передача нервного импульса может быть преобразована другими (возбуждающими или тормозными) нейронами.
Для вхождения в проблему активности нейрона в ЦНС (центральная нервная система) необходимо кратко остановиться на его морфологии. Мембрана нейрона. Нейроны обладают высоким уровнем морфологической и функциональной специализации, и первый уровень специализации проявляется в структуре и динамике его мембраны.
Как показывают результаты электронной микроскопии, плазматическая мембрана нейронов имеет такое же строение, что и у соматических клеток: она построена из липи¬дов и протеинов (цепочек аминокислот).
Основная структура мембраны двухслойная и представляет собой «сэндвич» из фосфолипидов, которые расположены таким образом, что полярные (заряженные) части прилежат к наружной части мембраны, а незаряженные части направлены вовнутрь клетки.
Такая организация максимизирует число гидрофобных и гидрофильных соединений, которые могут формировать и делать относительно прочной и очень тонкой оболочку, непроницаемую для большинства полярных молекул или ионов.
Мембрана является динамичной, и часто ее описывают как «жидко-мозаичную» структуру (Singer, Nicolson, 1972).
Липиды свободно диффундируют с одного участка на другой, обеспечивая тем самым мембране свойства жидкости.
Два жидких липидных слоя мембраны (наружный и внутренний) позволяют свободно плавать в ней специализированным бел¬кам и выполнять им свои функции.
Белки могут проникать через оба слоя, образуя каналы для транспорта через них ионов и небольших молекул.
Такие «интегральные белки» часто формируют межмембранные структуры.
Другие, «периферические белки», локализованы только в наружной или внутренней мембранах, они подвижны и выполняют определенные функции.
В жидкой мембране белки часто рассматриваются как частицы мембраны, произвольно плавающие в море липидов.
Следует отметить, что белки, проникающие через оба слоя мембраны, находясь одним концом снаружи, а другим внутри клетки, превращают участки мембраны в функциональные единицы, обеспечивающие определенные потребности нейрона.
Схема строения мембраны нейрона
Мембрана нейрона неоднородна, некоторые авторы выделяют в ней такие специфические участки, как дендритную зону с большим количеством синаптических контактов и пресинаптическую зону аксона. Кроме того, мембрана нейрона является асимметричной.
На ее наружной части (как и на мембране соматических клеток) находится большое количество карбогидратов сиаловой кислоты, которые обеспечивает отрицательный заряд наружной поверхности мембраны.
Белки мембраны.
По своей функции делятся на насосы, каналы, рецепторы, ферменты и структурные белки.
Насосы обеспечивают перемещение ионов и молекул против концентрационных градиентов и поддержание их необходимых концентраций в клетке.
Поскольку заряженные молекулы не могут пройти через двойной липидный слой, в мембранах есть набор специфических белковых канальцев, по которым вовнутрь клетки проходят определенные ионы.
Клеточные мембраны с помощью рецепторных белков узнают и прикрепляют к себе разные молекулы.
Ферменты размещаются внутри мембраны или на ней и облегчают протекание химических реакций у поверхности мембраны, например АТФаза, которая расщепляет АТФ — универсальную единицу топлива — для обеспечения теплом локальных химических процессов.
Структурные белки обеспечивают соединение клеток в органы и поддержание субклеточной структуры.
Не у всех мембранных белков функция жестко фиксирована.
Некоторые белки могут выполнять одновременно функции рецептора, фермента и насоса.
Кроме ионных насосов и канальцев, для выполнения основных функций нейронам требуются и другие белки.
Одним из таких белков является фермент аденилатциклаза, который регулирует внутриклеточную концентрацию циклического аденозинмонофосфата (циклического АМФ — цАМФ).
Циклические нуклеотиды, такие, как цАМФ, называют «вторичными мессенджерами». Внутри клетки цАМФ «собирает» информацию от первичных мессенджеров (нейромедиаторов) и подготавливает цитоплазму к возможным изменениям ее метаболизма.
Основная гипотеза заключается в том, что повышение концентрации цАМФ в ответ на поступление на постсинаптические рецепторы таких нейромедиаторов, как норадреналин и дофамин, ведет к повышению активности протеинкиназы, которая (1) фосфорилирует определенные белки мембраны и изменяет проницаемость мембраны, (2) изменяет метаболические процессы в клетке за счет активации и индукции некоторых ферментов и белков.
Общее заключение: цАМФ каким-то образом устанавливает уровень возбудимости нейрона (Cotman, McGaugh, 1980)
Понимание функций мембранных белков — один из этапов на пути к пониманию функций нейрона.
Подобно всем другим клеткам организма, в нейроне поддерживается постоянство внутренней среды, которая существенно отличается от окружающей нейрон межклеточной жидкости.
Особенно выражены различия в концентрациях ионов натрия и калия (Na*, Ka*). Наружная среда приблизительно в 10 раз богаче Na*, чем внутренняя, а внутренняя среда в 10 раз богаче Ka*, чем межклеточная жидкость. Данное различие в концентрации ионов натрия и калия лежит в основе поддержки и развития электрических потенциалов на мембране нервных клеток.
2. Ядро. В каждой нервной клетке есть ядро, в котором хранится генети¬ческий материал в виде хромосом. Хромосомы состоят из дезоксирибонуклеиновой кислоты (ДНК) и белков, которые вместе образуют гены.
Во время эмбрионального развития гены контролируют синтез белка и через белки обеспечивают дифференцировку клетки, ее ко¬нечную форму и синаптические связи с другими клетками.
В зрелом состоянии нейрона гены через контроль над синтезом белка контролируют активность нейрона.
Ядро отделено от цитоплазмы двумя мембранами, которые в некоторых местах сходятся и образуют поры, через которые осуществляется обмен веществ между цитоплазмой и содержимым ядра. 3. Митохондрии. Нейрон для выполнения своих функций нуждается в большом количестве энергии.
Макроэргическая молекула АТФ (аденозинтрифосфорная кислота) является основным источником энергии.
Подвижные и пластичные митохондрии представляют собой окруженные двойной мембраной органеллы, в которых осуществляется синтез АТФ.
Обычно митохондрии представляют собой мелкие (длиной 0,5-3 мкм) внутри¬клеточные образования, которые за счет своей подвижности располагаются в тех местах, где необходима энергия для поддержания химических процессов.
Для обеспечения внутриклеточных химических процессов теплом происходит расщепление (гидролиз) АТФ: от нее отделяется ион фосфора и энергия, которая удерживала этот ион с ионом АДФ (аденозиндифосфорная кислота).
Энергия идет на поддержание химических процессов в нейроне, а АДФ-3 и Р*3 поступают в митохондрии, в которых выделяющаяся от окисления глюкозы энергия идет на соединение этих ионов и образование АТФ.
Внутреннее содержимое тела клетки: Я— ядро, П — полисомы, Т— микротрубочки, М — митохондрии, МФ — микрофиламенты, Ш-ЭР — шероховатый эндоплазматический ретикулум.
4. Строительные белки-рибосомы, шероховатый эндоплазматический ретикулум и аппарат Гольджи
Эти структуры обеспечивают синтез белка в нейроне и его встраивание в структуры мембраны.
Белки нейронов, как и других клеток, нуждаются в постоянном обновлении.
Без обновления белков нейрон может прожить несколько дней.
В цитоплазме тела нейрона находится большое количество кластеров рибосом.
Рибосомы имеют размер около 4 нм в диаметре и сформированы из белков и рибонуклеиновой кислоты.
Кластеры рибосом, называемых полисомами, осуществляют в цитоплазме синтез растворимых белков, в том числе ферментов.
Отдельные рибосомы в полисомах связаны с информационной РНК (иРНК). иРНК является длинной цепочкой нуклеиновых кислот, представленных четырьмя нуклеотидами: аденином, гуанином, цитозином и урацилом.
Последовательность этой нуклеотидной цепочки кодирует последовательность аминокислот в синтезируемом белке.
Специальная транспортная РНК (тРНК) «распознает» определенную тринуклеотидную цепочку на матричной РНК (мРНК) и связывает с ней определенную аминокислоту.
По мере синтеза белка мРНК продвигается через рибосому и к ее тринуклеотидным цепочкам тРНК последовательно присоединяют разные аминокислоты до тех пор, пока мРНК не закончится.
Затем цепочка аминокислот выходит в цитоплазму (Ленинджер, 1982)
Синтез мембранных белков и их включение в мембраны осуществляется с помощью шероховатого поверхностного эндоплазматического ретикулума (шероховатого ЭР), гладкого эндоплазматического ретикулума (гладкого ЭР) и аппарата Гольджи.
Шероховатый ЭР представляет собой лабиринтную систему мембранных трубок, пузырьков и цистерн, чья выступающая вовнутрь нейрона поверхность усыпана рибосомами, связанными друг с другом с помощью мРНК.
Отсюда и термин «шероховатая поверхность».
Мембранные белки внедряются в шероховатый ЭР.
Гладкий ЭР является продолжением шероховатого ЭР и лишен рибосом.
Гладкий ЭР вовлечен в распределение белка по нейрону: а именно по нему вновь синтезированные белки доставляются в дендриты.
Из гладкого ЭР белки транспортируются в аппарат Гольджи, где они могут быть модифицированы, например если этот белок относится к гликопротеинам, то к нему добавляется карбогидрат.
Белки в аппарате Гольджи концентрируются и затем «упаковываются» в мембранных пузырьках и изолируются для последующей поставки его в другие участки клетки.
По соседству с аппаратом Гольджи лежат «облачки» мелких пузырьков, которые, возможно, транспортируют синтезированные белки в разные участки нейрона. Лизосомы. Лизосомы относятся к внутриклеточной пищеварительной системе. Эта структура, как и ретикулум, заключена в мембрану.
Лизосомы не имеют определенной формы или размера.
Они содержат разнообразные гидролитические ферменты, которые расщепляют и переваривают множество соединений, появляющихся как внутри, так и вне клетки.
Перевариваемые вещества могут быть внутриклеточными, и такое переваривание называют аутофагией. Переваривание лизосомами внеклеточных веществ получило название гетерофагия. Цитоскелетная сеть.
В теле и отростках нейрона имеется обширная цитоскелетная сеть, состоящая из микротрубочек, нейрофиламентов и микрофиламентов.
Они проходят через весь нейрон, соединяя все его части.
Эта сеть является каркасом нейрона, поддерживая определенную его форму.
С другой стороны, цитоскелетная сеть выполняет транспортную функцию.
В теле клетки микротрубочки и более мелкие трубочки, нейрофиламенты и микрофиламенты, занимают большую ее часть, не занятую другими органеллами, и из тела нейрона все эти трубочки проникают в дендриты и аксоны.
Микротрубочки состоят из длинных неразветвленных трубочек разной длины.
Стенки их построены из субъединиц специфического белка — тубулина (от лат. tubula — трубочка).
Нейрофиламенты тоньше микротрубочек.
Они тоже имеют трубчатое строение и встречаются только в нейронах.
Показано, что в крупных аксонах их значительно больше, чем микротрубочек, тогда как в мелких аксонах и дендритах их соотношение противоположное.
Нейрофиламенты и их соотношение с микротрубочками меняются при старении.
При болезни Альцгеймера они превращаются в клубочки и бляшки.
Предполагается, что микротрубочки и нейрофиламенты в аксонах и дендритах выполняют транспортную функцию между телом и отростками нейрона в обоих направлениях: от тела к отросткам — антероградный транспорт, от отростков к телу — ретроградный транспорт.
Данная гипотеза была подтверждена экспериментально.
После инъекции меченых аминокислот вблизи тела клетки методом радиографии было показано, что эти аминокислоты поглощаются телами нейронов и включаются в белок, который затем переносится по аксону и его коллатералям.
В этих экспериментах были выявлены два типа аксонного транспорта: медленный транспорт, идущий со скоростью 1 мм в сутки, и быстрый, идущий со скоростью несколько сотен миллиметров в сутки.
Многие переносимые вещества связаны с функциями синаптической передачи.
Микрофиламенты присутствуют в большом количестве в нервных отростках.
Их много в нейроглии, и они участвуют в некоторых связях между нейронами (Шеперд, 1997).
Дендриты.
Дендриты при всем их разнообразии среди нейронов содержат те же органеллы, кроме ядра, что и тело.
В большей части дендритов параллельно проходит большое количество микротрубочек.
С другой стороны, в дендритах находится лишь небольшое количество нейрофиламентов.
Митохондрии ориентированы вдоль дендрита и достигают по длине 9 мкм.
Шероховатый ЭР хорошо выражен лишь в начале дендрита; по мере удаления его от тела ЭР исчезает.
Гладкий ЭР распространен по всей длине дендритов.
Он проходит параллельно микротрубочкам и микрофиламентам, образуя по своему курсу небольшие выпячивания.
Считается, что гладкий ЭР распространен по всему дендриту и транспортирует разные химически вещества.
Основная часть белка синтезируется в теле клетки, но некоторые белки синтезируются в проксимальных участках дендритов, где локализована большая масса гладкого ЭР.
Эти белки транспортируются в дистальные отделы дендритов с помощью цистерночек и гладкого ЭР вдоль поверхности системы микротрубочек.
Белки, которые синтезируются в теле клетки, транспортируются в дендриты также с помощью гладкого ЭР.
На дендритах располагается большое количество шипиков, на ко¬торых находятся в основном аксодендритные синапсы.
Например, на дендритах пирамидных нейронов в среднем локализовано около 4000 шипиков, что составляет примерно 43% от всей синаптических контактов этих нейронов (Р (Cotman, McGaugh, 1980).
Каждый шипик представляет собой выпячивание на дендрите длиной около 2 мкм, которое состоит из тонкой шейки, заканчивающейся яйцеобразным выпячиванием.
Цитоплазма шипиков заполнена тонкими филаментами и очень небольшим количеством микротрубочек. Как было сказано выше, шипики включены в синаптические структуры.
Аксон. В отличие от дендритов аксон является обычно одиночным отростком.
В нем нет шероховатого ЭР, рибосом, но он содержит митохондрии, большое количество нейрофиламентов, микротрубочки, гладкий ЭР и небольшое количество лизосом.
Часть гладкого ЭР, цистерны разной формы, соединены друг с другом тонкими трубочками, и эта система идет вдоль всего аксона.
Внутри аксон заполнен желеобразной аксоплазмой, которая удерживает его цитоскелетную сеть.
У основного большинства крупных нейронов аксоны на всем протяжении покрыты оболочкой, называемой миелином.
Миелин никогда не покрывает дендриты.
Известно, что миелин производится из плазматической мембраны глиальных клеток, которые обматывают аксон.
В периферической нервной системе аксоны обматывают особые глиальные, или Шванновские клетки.
Оголенные участки аксона между Шванновскими клетками называются перехватами Ранвье.
В ЦНС миелиновые оболочки формируются олигодендроцитами.
По миелинизированным аксонам потенциалы действия проходят быстро за счет сальтаторного (прыжкообразного) перемещения от одного перехвата Ранвье к другому. Синапс. Нервно-мышечное соединение.
Все синапсы имеют одинаковую структуру.
Пресинаптическая и постсинаптическая мембраны являются высокоспециализированными в месте контакта и образуют синаптическое соединение.
Пресинаптическая бляшка, или терминаль аксона, содержит пузырьки диаметром около 20-65 нм, которые наполнены веществом — нейромедиатором.
Наличие пузырьков и синаптического соединения является показате¬лем химического синапса.
Общие детали строения едины для всех синапсов, но тонкая структура синапсов зависит от особенностей пре- и постсинаптического нейронов, эта структура существенно различна в нервно-мышечном и межнейронных синапсах.
Нервно-мышечное соединение: А — аксон, Б — синаптические окончания аксона в виде бляшек, М — мышца, мт — митохондрии, сп — синаптические пузырьки, а — активная зона, Щ — синаптическая щель, сс — соединительные складки мембраны мышечной клетки.
Непосредственно перед нейромускулярным соединением миелиновая оболочка на аксоне исчезает, и далее аксон, окруженный только тонким слоем цитоплазмы Швановских клеток, разделяется на веточки (коллатерали).
В месте синаптического контакта коллатерали аксона с мышечной клеткой мембрана последней образует множе¬ство складок — соединительные складки.
Пространство между пресинаптической и постинаптической частями синапса получило название синаптической щели. Синаптическая щель в нейромускулярном соединении шире, чем между нейронами ЦНС.
В синаптических окончаниях аксонов имеется несколько митохондрий, сеть фиброзного белка, и множество синаптических пузырьков, содержащих нейромедиатор, а именно до 10000 молекул ацетилхолина, который вызывает возбуждающее действие на мембране мышечной клетки.
Некоторые пузырьки распределены в случайном порядке по аксонной ветви, в то время как другие находятся вдоль пресинаптического участка — активной зоны, из которой они выделяются в синаптическую щель.
Внутри двухслойной плазматической мембраны в активной зоне находятся внутримембранные частицы с участками, к которым крепятся пузырьки.
При прохождении по аксону потенциала действия пузырьки подплывают к этим участкам и сливаются с ними.
Напротив активной зоны пресинаптической мембраны находятся функциональные складки мембраны мышечной клетки.
Внутри мембраны, в основном в гребнях складок, имеется большое количе¬ство внутримембранных включений, с которыми, по-видимому, соединяется нейромедиатор. Синапсы ЦНС. Каждый синапс в ЦНС сформирован двумя нейронами — пре- и постсинаптическим.
Как и в нейромускулярном соединении, в этих синапсах есть пре- и постсинаптическая мембраны, между которыми имеется синаптическая щель шириной порядка 20-50 нм.
В пресинапсе находятся пузырьки с нейромедиатором, митохондрии, мембранные цистерночки, случайные микротрубочки и множество волокон.
В ЦНС выделяют два типа синапсов.
Синапсы ЦНС отличаются от синапсов нервно-мышечного соединения в нескольких аспектах.
Если в нервномышечном соединении ацетилхолин оказывает возбуждающее действие на мембрану мышечной клетки, то в ЦНС в одних межнейронных синапсах ацетилхолин оказывает возбуждающее действие на мембрану нейрона, в других синапсах — тормозное действие.
Если в нервномышечных соединениях существует только один нейромедиатор, то в синапсах ЦНС насчитывается более 100 различных нейромедиаторов.
Первый тип синапсов имеет довольно типичную форму.
Пресинаптическая мембрана аксона имеет множество уплотненных проекций на ее цитоплазматическую часть, а постсинаптическая мембрана дендрита также имеет уплотнения, но непрерывные, на цитоплазматической поверхности — постсинаптическое уплотнение. Типичная длина такого синаптического контакта примерно 5 мк. Синапсы 1-го типа являются самыми многочисленными; они всегда являются аксо- дендритными, формируются на шипиках дендрита и никогда не формируются на теле нейрона.
Уплотненные проекции пресинаптической мембраны представляют собой серии пирамид, организованных в гексагональные структуры . Активные зоны синапсов находятся в промежутках между этими структурами. В активных зонах находятся синаптические пузырьки, а также множество внутримембранных включений.
Постсинаптические уплотнения представляют собой фиброзную ткань, напоминающую грубо сотканный ковер.
Эта ткань лежит напротив постсинаптической мембраны.
В этих уплотнениях срастаются тонкие волокна, к которым прикасаются микротрубочки, содержащие включения небольшого размера.
Наружная поверхность постсинаптической мембраны, перекрывающая данное уплотнение, является чувствительной к определенному нейромедиатору; множество щетинок и волокон ориентированы в направлении синаптической щели, и некоторые из них соединяются с пресинаптической мембраной.
Второй тип синапсов никогда не формируется на шипиках.
Они обычно локализованы на телах нейронов.
Предполагается, что в отличие от синапсов 1-го типа синапсы 2-го типа являются тормозными синапсами. Основной признак 2-го типа синапсов — это отсутствие выпячивающей постсинаптической плотности; область плазматической мембраны в синапсе очень тонкая.
Искусственная стимуляция нервной клетки[править | править код]
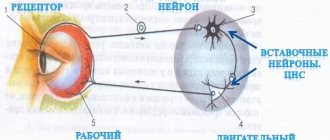
Когда электрический импульс из внешнего источника приложен к нервной клетке, ток течет от положительно заряженного электрода (анода) и выходит на отрицательно заряженный электрод [катод). Нервное волокно ниже катода деполяризуется, и при условии, что достигнут пороговый потенциал, генерируется потенциал действия.
Скорость проведения импульса по нерву можно измерить, поместив два электрода на кожу по ходу нерва на известном расстоянии друг от друга, с последующей стимуляцией этого нерва (содержащего многочисленные нейроны) и регистрацией времени, которое потребовалось суммарному потенциалу действия для прохождения расстояния между электродами. Скорость проведения сигнала у человека обычно составляет от 40 до 70 м/с. Значения ниже 40 м/с считаются патологическими.
- Случайное воздействие электричества. Высокое напряжение, особенно низкочастотный переменный ток (например, при контакте с электрической розеткой), а также в условиях сниженного сопротивления (босые ноги, несчастный случай в ванной), воздействует в основном на проведение сигналов в сердце, что может вызвать фибрилляцию желудочков.
Постоянный ток обычно действует как стимул только при включении и выключении: высокочастотный переменный ток (> 15 кГц), напротив, не способен вызвать деполяризацию, но повреждает ткани организма. На этом принципе основана диатермия.