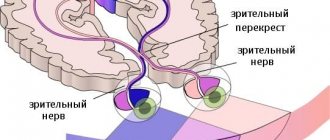
Centripetal fibers
Afferent nerve pathways are classified into unconscious and conscious sensory pathways. It is with their help that communication is ensured between all integration centers located in the brain. For example, they provide a direct connection between the cerebellum and the cerebral cortex.
The main afferent pathways of the central nervous system for conscious general sensitivity are fibers of pain, temperature and tactile sensitivity, as well as conscious proprioceptive. The main unconscious pathways of general sensitivity are the anterior and posterior spinocerebellar. Special conductors include vestibular, auditory, gustatory, olfactory and visual.
SENSE ORGANS
Some patterns of the structure of efferent projection pathways
1. The first neuron of all efferent pathways is localized in the cerebral cortex.
2. Efferent projection pathways occupy the anterior peduncle, knee and anterior part of the posterior limb of the internal capsule, passing at the base of the cerebral peduncles and pons.
3. All efferent pathways end in the nuclei of the motor cranial nerves and in the anterior horns of the spinal cord, where the last motor neuron is located.
4. Efferent pathways form a complete or partial decussation, as a result of which impulses from the cerebral cortex are transmitted to the muscles of the opposite half of the body.
The sense organs perceive various irritations acting on the human and animal body, as well as the primary analysis of these irritations. Academician I.P. Pavlov defined the senses as peripheral zones of analyzers. Their specific perceiving elements are sensitive nerve endings - receptors that convert the energy of an external stimulus into nerve impulses. The latter contain in encoded form information about objects and phenomena of the external world. These impulses are transmitted along afferent nerve pathways to the subcortical and cortical centers, where the final analysis of stimulation occurs. According to the doctrine of analyzers, afferent pathways represent their middle, conductive section, and the perceptive zones of the cortex are their central ends. The emergence of sensations is associated with the cortical sections of the analyzers.
In protozoa, sensitivity is inherent in the outer layer of protoplasm of their single cell. In lower animals, whose body consists of endoderm and ectoderm, all cells of the latter respond to external stimuli. Simultaneously with the differentiation of the muscular and nervous systems, individual perceptive cells are isolated in the ectoderm, which are associated with the central nervous system and represent the primary sensory cells: they are initially (in lower coelenterates) scattered throughout the body, then grouped in certain places, especially in the circumference of the mouth. Such groups of sensory cells are the simplest sensory organs in structure and function. Finally, more advanced forms are observed in higher ones, where the sensory organs include not only perceptual elements, but also special additional (auxiliary) devices: first, indifferent (supporting) epithelial cells, then connective and muscle tissue.
In the process of evolution, organs develop that are adapted to perceive a wide variety of environmental agents - mechanical, physical, chemical. For example, termites perceive a magnetic field, bees and ants - ultraviolet rays, cockroaches and squids - infrared rays, fish have a lateral line organ that perceives the direction and speed of water movement, and shrews and bats are capable of perceiving ultrasonic vibrations. In higher animals and humans, the sense organs are the organ of smell, the organ of taste, the organ of vision, the vestibulocochlear organ and the skin, which, together with its appendages, forms the general covering of the body.
Based on the characteristics of development, structure and function, 3 types of sensory organs are distinguished. Type I includes the organs of vision and smell, which are formed in the embryo as part of the brain. Their structure is based on primary sensory, or neurosensory, cells. These cells have specialized peripheral processes that perceive vibrations of light waves or molecules of volatile substances, and central processes through which excitation is transmitted to afferent neurons.
Type II includes the organ of taste, hearing and balance. They are formed in the embryonic period in the form of thickening of the ectoderm and placodes. Their main receptor element is secondary sensory sensory epithelial cells. Unlike neurosensory cells, they do not have axon-like processes. The excitation that occurs in them under the influence of flavoring substances, vibrations of air or a liquid medium is transmitted to the endings of the corresponding nerves.
The third type of sensory organs is represented by receptor encapsulated or non-encapsulated bodies and formations. These include receptors in the skin and subcutaneous tissue. They are nerve endings surrounded by connective tissue or glial cells. A common feature of all perceptive cells is the presence of flagella - kinocilia or microvilli - stereocilia. Molecules of special photo-, chemo- and mechanoreceptor proteins are embedded in the plasma membrane of flagella and microvilli. These molecules perceive influences of only one specific type and encode them into specific cell information, which is transmitted to the corresponding nerve centers.
Sense organs also differ in the complexity of their anatomical structure. The organs of taste and skin sense, which are represented mainly by epithelial formations, are relatively simple in structure. The organs of smell, vision, hearing and balance have auxiliary devices that ensure that they receive only those irritations to which these sense organs are adapted to perceive. Thus, the auxiliary apparatus of the olfactory organ is the ethmoid labyrinth and paranasal sinuses, which direct the air stream to the olfactory receptors. The organ of vision is equipped with an optical apparatus that casts images of external objects onto the retina of the eye. The organ of hearing has a complex apparatus for capturing and conducting sounds.
Auxiliary apparatus of the sensory organs not only ensure the interaction of specific stimuli with receptors, but also block the path of extraneous, inappropriate stimuli, and also provide protection for the sensory organs from external mechanical influences and damage.
Fibers of tactile, temperature and pain sensitivity
This path originates from receptors in the epithelium, impulses from which enter the cells of the spinal ganglion, and then into the spinal cord, to the nuclei of the optic thalamus. Then to the cortex of the postcentral gyrus, where their complete analysis takes place. Three tracts are involved in this pathway:
- Thalamo-cortical.
- Gangliospinal.
- The lateral spinothalamic tract, which runs in the lateral funiculus of the spinal cord and the tegmentum of the brain stem.
The trigeminal nerve is responsible for receiving tactile sensations in the facial part of the head and changes in body temperature. When it is damaged, a person begins to experience severe pain in the facial area, which either disappears or reappears. The trigeminal nerve passes through the cervical spine where the motor fibers of the corticospinal tract cross. The axons of the sensory neurons of the trigeminal nerve pass through one of the parts of the medulla oblongata. Through these axons, the brain receives information about pain in the mouth, teeth, and also in the upper and lower jaws.
Projection and association areas of the cortex
text_fields
text_fields
arrow_upward
The bodies of afferent neurons, as a rule, lie in the sensory spinal or cranial ganglia. The exception is the visual and olfactory systems, where sensory neurons are located directly in the retina (ganglion cells) or olfactory bulb, respectively. The processes of these neurons enter the spinal cord or brain, where a switch to the next-order neuron occurs. Further along the network of neurons the signal propagates in an upward direction. For most sensory systems, except for the olfactory system, the penultimate neuron lies in specific nuclei of the thalamus. From here, information enters the corresponding projection and associative zones of the cortex, where memory processes are included in signal processing. It was noted that impulses from receptors reach the primary projection zones of the cortex via the shortest route, while activation of associative zones occurs somewhat later due to the involvement of polysynaptic nerve networks in this process (see Fig. 3.45).
Rice. 3.45.
Rice.
3.45. The system of connections between the fields of the human cerebral cortex (according to Polyakov): I - primary (central) fields; II – secondary (peripheral) fields; III - tertiary (associative) fields (overlap zones of analyzers). Bold lines highlight: the system of projection (cortical-subcortical) connections of the cortex; system of projection-associative connections of the cortex; system of associative connections of the cortex. 1 - receptor; 2 – effector; 3 – sensory ganglion neuron; 4 – motor neuron; 5–6 – switching neurons of the spinal cord and brainstem; 7–10 – switching neurons of subcortical formations; 11, 14 – afferent fiber from the subcortex; 13 – pyramid of layer V; 16 and 18 – pyramids of layer III; 12, 15, 17 – stellate cells of the cortex.
Fibers of conscious general sensitivity
This path carries through itself all types of general sensitivity from the head to the neck. The receptors begin their journey in the muscles and skin, conduct impulses to the sensory ganglia and pass into the nuclei of the trigeminal nerve. Next, the path passes to the visual thalamus, and then spreads to the cells of the postcentral gyrus. This includes three main paths:
- thalamocortical;
- ganglionuclear;
- nuclear-thalamic.
Fibers of conscious proprioceptive sensitivity
This pathway originates with its receptors in the tendons, periosteum, muscles and ligaments, as well as in the joint capsules. In this case, complete information is provided about vibrations, body position, degree of relaxation and muscle contraction, pressure and weight. The neurons of this pathway are located in the spinal ganglia, the nuclei of the sphenoid and thin tubercles of the medulla oblongata, and the optic thalamus of the diencephalon, in which the switching of impulses then begins. Information is analyzed and ends its path in the central gyrus of the cerebral cortex. This path includes three tracts:
- Thalamocortical, which ends in the projection center, that is, in the central gyrus of the brain.
- Thin and wedge-shaped bundles passing in the posterior cord of the spinal cord.
- The bulbar-thalamic tract passes through the tegmentum of the brain stem.
SENSITIVE (afferent) CONDUCTING PATHWAYS
IV. pathways of the brain and spinal cord
Pathways are a chain of neurons connecting functionally homogeneous areas of gray matter in the central nervous system, occupying a certain place in the white and gray matter of the brain and spinal cord and conducting the same impulse.
The pathways are part of complex reflex arcs that connect various parts of the central nervous system and provide two-way functional communication between individual structures of the brain and spinal cord. They are distinguished by their number, complexity of structure and reliability of operation.
Depending on the size, shape and direction of the nerve impulse, the conducting pathways are named: path (tractus),
bundle
(fasciculus),
fiber
(fibrae),
commissure
(commissura),
loop
(lemniscus)
or radiance
(radiatio).
All pathways of the central nervous system are divided into three groups:
1. Projection.
2. Commissural.
3. Associative.
In the process of formation of the conduction system in ontogenesis, projection pathways are initially formed, and then commissural and associative ones.
Associative pathways connect parts of the cortex within one hemisphere.
There are:
A. Short fibers, arcuate fibers, fibrae arcuatae cerebri,
which connect the cortical fields of neighboring gyri.
B. Long fibers that connect the cortical fields of distant gyri (superior longitudinal fasciculus, fasciculus longitudinalis superior,
lower longitudinal fascicle,
fasciculus longitudinalis inferior
, belt,
cingulum
, uncinate fascicle,
fasciculus uncinatus
).
Commissural pathways connect the symmetrical parts of the right and left hemispheres. These include:
1. Corpus callosum, corpus callosum.
2. Anterior commissure, commissura anterior.
Refers to the olfactory brain.
3. Commissura of the fornix, commissura hippocampi (fornicis).
Connects the cortical fields of the hippocampus of the right and left hemispheres.
4. Posterior commissure, comissura posterior.
5. Adhesion of leashes, comissura habenulare
.
6. Interthalamic fusion, adhesio intertalamica
.
Projection pathways connect the cortex with the subcortical formations of the brain and the spinal cord.
Projection pathways connect the cerebral cortex with its underlying sections (short pathways) and with the spinal cord (long pathways).
Based on the direction of nerve impulse conduction, projection pathways are divided into two groups:
• afferent (ascending, centripetal, sensitive), which conduct nerve impulses from receptors that perceive information from the external world or the internal environment of the body to various parts of the brain and to the cerebral cortex;
• efferent (descending, centrifugal, motor), transmitting impulses from the cerebral cortex and other parts of the brain to the periphery.
SENSITIVE (afferent) CONDUCTING PATHWAYS
Sensitivity is understood as the body’s ability to perceive the effects of stimuli from the external and internal environment.
The type of sensitivity is determined by the nature of the receptor that perceives the irritation. Among the receptors there are exteroceptors (tactile, pain, temperature), located in the skin and mucous membranes.; proprioceptors (musculo-articular, vibration, sense of pressure and weight), which are located in muscles, tendons, ligaments, joint capsules; and visceroreceptors (sensitivity of internal organs and blood vessels), located in various internal organs and vessels.
Depending on the type of sensitivity, afferent pathways are divided into:
• exteroceptive (contact – general sensitivity and distant – specific sensitivity,
• proprioceptive (cortical – conscious and cerebellar – unconscious),
• interoceptive.
Those of them that end in the cerebral cortex are called conductors of conscious sensitivity, or cortical pathways.
Sensitive pathways of the cortical direction are characterized by:
1. Presence of 3 neurons.
2. The first neurons are represented by pseudounipolar neurons, the bodies of which are located in the spinal ganglion, and the peripheral processes form receptors.
3. The bodies of II neurons are located in the nuclei of the spinal cord or medulla oblongata.
4. Axons of II neurons form (as a rule) a decussation.
5. Most of the axons of II neurons go to the optic thalamus as part of the medial lemniscus.
6. The fibers of these pathways are located, as a rule, in the dorsal parts of the brain stem.
7. The bodies of III neurons are located in the lateral parts of the visual thalamus.
8. All of them pass in the middle third of the posterior thigh of the internal capsule.
9. Axons of III neurons on the way to the cortex form or pass as part of the corona radiata.
Spinal fibers
The afferent pathways of the spinal cord are formed with the help of axons, or, as they are also called differently, the endings of neurons. Axons are located only in the spinal cord and do not extend beyond it, and also create connections between all segments of the organ. The atomic structure of these fibers is that the length of the axons is quite large and connects to other nerve endings. Nerve signals are transmitted from the receptors to the central nervous system through the afferent pathways of the spinal cord and brain. All nerve fibers located along the entire length of the spinal cord are involved in this process. The signal to the organs is carried out from different parts of the central nervous system and between neurons. The unimpeded passage of signals from the periphery to the central nervous system is achieved using the spinal cord pathways.
Posterior and anterior spinocerebellar tracts
The afferent pathways of the cerebellum are unconscious and originate in the lateral cord of the spinal cord, and from there they carry information about the state of the organs of the musculoskeletal system. The anterior spinocerebellar tract enters the cerebellum through the superior peduncle, and therefore passes through the tegmentum of the medulla oblongata, midbrain and pons. The posterior spinocerebellar tract runs in the medulla oblongata and enters through the inferior peduncle.
These two tracts transmit information from ligaments, joint capsules, muscle receptors, tendons, and periosteum to the cerebellum. They are responsible for maintaining balance and coordinating human movements, so their role in the body is very important.
Auditory fibers
This pathway carries information from the receptors of the organ of Corti, which is located in the inner ear. Nerve impulses enter the bridge, which contains the auditory nuclei, along the fibers of the vestibulocochlear nerve. Through the auditory nuclei, information is transmitted to the nuclei of the trapezoid body. After this, the impulses arrive at the subcortical hearing centers, which include the thalamus optic, inferior colliculi and medial geniculate bodies.
In the midbrain, return reactions arise to these auditory stimuli, while the afferent auditory pathways switch to the nuclei of the thalamus, in which auditory stimuli are evaluated - they are responsible for movements that occur involuntarily: walking, running. Auditory radiation begins to emanate from the geniculate bodies - this tract conducts impulses from the internal capsule to the projection center of hearing. Only here does the evaluation of sounds begin to take place. The associative auditory center is located in the posterior part of the temporal gyrus. It is in it that all sounds begin to be perceived as words.
Some patterns of the structure of afferent
Projective pathways.
1. The beginning of each path is represented by receptors located in the skin, subcutaneous tissue or deep parts of the body.
2. The first neuron of all afferent pathways is located outside the central nervous system, in the spinal ganglia.
3. The second neuron is localized in the nuclei of the spinal cord or medulla oblongata.
4. All ascending pathways pass through the tegmentum of the brainstem.
5. The third neuron in the pathways heading to the cerebral cortex is located in the nuclei of the thalamus, and in the cerebellar pathways - in the cerebellar cortex.
6. The pathways bringing impulses to the cerebral cortex have one crossing, made by the processes of the 2nd neuron; thanks to this, each half of the body is projected onto the opposite hemisphere of the cerebrum.
7. The cerebellar tracts either do not cross at all, or cross twice, so that each half of the body is projected onto the cortex of the same half of the cerebellum.
8. The pathways connecting the cerebellum to the cerebral cortex are crossed.
Efferent pathways.
Descending projection pathways (effector, efferent) conduct impulses from the cortex, subcortical centers to the underlying sections, to the nuclei of the brain stem and motor nuclei of the spinal cord. These pathways are divided into 2 groups: 1) pyramidal pathway and 2) extrapyramidal pathways.
Pyramid Path
Pyramid path (tractus pyramidalis)
connects neurons of the motor cortex directly with the motor nuclei of the spinal cord and cranial nerves. The beginning of the path is large pyramidal neurons (Betz cells) (1st neuron), located in the inner pyramidal layer of the cortex of the precentral gyrus (primary cortical area 4).
The pyramidal tract is divided into the corticospinal tract (fibrae corticospinales)
and
corticonuclear (fibrae corticonucleares)
. The latter will be considered when describing the pathways of cranial nerves.
The corticospinal tract is composed of axons of large pyramidal neurons located in the upper and middle thirds of the precentral gyrus. They pass through the anterior part of the posterior limb of the internal capsule, the middle part of the base of the cerebral peduncle, the base of the pons and the pyramid of the medulla oblongata. At the border with the spinal cord there is an incomplete crossover of the corticospinal tract (decussatio pyramidum)
.
Most of the fibers, passing to the opposite side, form the lateral corticospinal tract (tractus corticospinalis lateralis)
, the rest go to
Rice. 7. Pyramid path.
composition of the anterior corticospinal tract (tractus corticospinalis anterior)
on their side and intersect in the spinal cord segment by segment, passing through the white commissure. They predominantly end in the intermediate gray matter, forming synapses with interneurons (2nd neuron), which transmit impulses to the motor neurons of the anterior horn nuclei (3rd neuron). The axons of the motor cells of the anterior horns exit the spinal cord in the anterior roots and then go as part of the spinal nerves to the skeletal muscles, providing their motor innervation.
Through intercalary neurocytes, the cells of the cerebral cortex communicate with the motor neurons that innervate the muscles of the neck, trunk and proximal limbs. The majority of the fibers of the lateral pyramidal tract end on the intercalated neurocytes of the spinal cord.
Motor neurons of the spinal cord, innervating the muscles of the forearm and hand, have direct connections with the cells of the cortex. Corticospinal fibers end on them (and not on interneurons), passing mainly as part of the anterior pyramidal tract. This achieves direct, more advanced cortical regulation of muscles, which are characterized by very precise, strictly differentiated movements.
The pyramidal tract primarily transmits signals to muscles for voluntary movements, regulated by the cerebral cortex. When it is interrupted, paralysis of the muscles of one's own or the opposite side occurs, depending on the level of damage. When the pyramidal tracts are damaged, the finely differentiated movements of the upper extremities, the muscles of which have cross innervation, are especially affected. The muscles of the lower extremities and especially the muscles of the trunk are innervated along with crossed and also uncrossed corticospinal fibers, so their function is impaired to a lesser extent.
However, the pyramidal tract connects the cortex not only with motor neurocytes of the spinal cord, but also with other structures of the brain and spinal cord. Its fibers or their collaterals end on the cells of the red nucleus, the pons own nuclei, and the cells of the reticular formation of the brain stem.
Extrapyramidal tracts.
The extrapyramidal system is represented by multi-link descending pathways, through which the regulation of involuntary movements, automatic motor acts, muscle tone, as well as movements expressing emotions (smile, laugh, cry, etc.) is carried out.
Neurons of the inner pyramidal layer of the frontal lobe cortex (field 6) (1-neuron) send corticostriatal fibers to the new part of the striatum, represented by the caudate nucleus and putamen. The 2nd neuron of the extrapyramidal tract is localized here, the processes of which go to the ancient part of the striatum - the globus pallidus (striato-pallidal fibers). The nerve cells of the globus pallidus represent the 3rd neuron, their axons run as part of the lenticular loop (ansa lenticularis)
to various nuclei of the brain stem - subthalamic nucleus, substantia nigra, nuclei of the superior colliculus, red nucleus, lateral vestibular nucleus,
Rice. 9. Cortico-pontine-cerebellar-spinal tract.
olivary nucleus, reticular nuclei. These nuclei contain the 4th neuron, which gives rise to descending pathways that transmit signals to the motor nuclei of the cranial nerves and spinal cord, the tectospinal cord (tractus tectospinalis),
the red nucleus-spinal cord (tractus vestibulospinalis), olivo -spinal cord (tractus vestibulospinalis). spinal (tractus olivospinalis), reticular-spinal (fasciculi reticulospinales)
. The most well developed in humans are the red nucleus-spinal cord and reticular-spinal cord tracts, the rest are represented by a few fibers and are usually traced only in the cervical segments of the spinal cord. The motor cells of the nuclei of the cranial nerves and the anterior horns of the spinal cord form the 5th neuron of the extrapyramidal tract, which sends impulses to the skeletal muscles.
The extrapyramidal tract also includes a system of fibers connecting the cerebral cortex with the cerebellum. This system consists of two links. The first is the cortical-pontine tract (tractus corticopontini)
.
They originate from the cells of the inner pyramidal layer of the secondary cortical fields of the frontal (fields 6 and 8), parietal, occipital, and temporal lobes; Accordingly, the frontal, parietal, occipital and temporopontine pathways are distinguished. All of them pass through the anterior leg of the internal capsule, the base of the cerebral peduncle (its medial and lateral sections) and end in the own nuclei of the bridge, where the 2nd neuron (nuclei pontis)
.
The axons of the cells forming the nuclei of the bridge pass along the middle cerebellar peduncles as part of the tractus pontocerebellaris
to the opposite side (1st chiasm) and end in the cortex of the cerebellar hemispheres (new cerebellum).
The piriform cells of the cerebellar cortex are taken to be the 3rd neuron. The impulses they send enter the dentate nucleus (4th neuron). From here, impulses are transmitted along the dentate-red nuclear pathway (tractus dentatorubralis)
through the superior cerebellar peduncles to the red nucleus (5th neuron).
The 2nd decussation (decussatio pedunculorum cerebellarium superiorum)
.
From the red nucleus begins the red nucleus-spinal tract (tractus rubrospinalis)
, which after the decussation
(decussatio ventralis tegmenti)
goes to the nuclei of the anterior horns of the spinal cord and the motor nuclei of the cranial nerves (6th neuron). From here, as part of the spinal and cranial nerves, impulses enter the muscles.
Rice. 8. Extrapyramidal tracts.
Through the corticopontine and pontocerebellar pathways and the ascending efferent pathways of the cerebellum, a circular interaction is carried out between the cerebral cortex and the cerebellum, which is necessary for the regulation and coordination of various motor acts. The cerebellum receives from the cerebral cortex, as it were, copies of commands sent along the pyramidal and extrapyramidal pathways, compares them with signaling coming from the proprioceptors and vestibular apparatus, and sends the processed information to the higher motor centers of the cortex.
Taste analyzers
The impulses of the afferent pathway of taste analyzers develop from the receptors of the root of the tongue, which are part of the glossopharyngeal nerves and located on the tongue, which are part of the facial nerve. Impulses from them enter the medulla oblongata, and then to the nuclei of the facial and glossopharyngeal nerve. The smallest part of all information received from these impulses is delivered to the cerebellum, thereby forming the nuclear-cerebellar pathway, and provides reflex regulation of the tone of the muscles of the tongue, head and pharynx. Most of the information enters the visual thalamus, after which the impulses reach the uncinus of the temporal lobe, where they are consciously analyzed.
Lesya Ukrainka
- pseudonym of Larisa Petrovna Kosach, a famous Ukrainian writer and poet.
Lesya Ukrainka is a consistent and energetic fighter for the education of the Ukrainian people, for their consolidation into a political nation. The entirety of her works is an image of an ideal earth, an earthly paradise called.
Lesya Ukrainka was born on February 25, 1871 in Novograd-Volynsky
, in that part of Ukraine that was part of the Russian Empire.
The family is not alien to high spiritual interests: the mother
is a writer who wrote under the pseudonym Olena Pchilka (her poetry and stories in her native language for children were well known in Ukraine),
the father
is a highly educated landowner who was very fond of literature and painting. Writers, artists and musicians often gathered in the Kosachs’ house, and evenings and home concerts were held. Uncle Lesya (that was her name in the family, and this home name became her literary pseudonym.) - Mikhail Drahomanov, who later took friendly care of his niece and helped her in every way - was a famous scientist, public figure, lived for a long time abroad in France and Bulgaria. He made acquaintance with Ivan Sergeevich Turgenev, Victor Hugo, was aware of all the latest literary and political events and often replenished his niece’s library with parcels from abroad.
Lesya Ukrainka began writing and publishing as a child (12 years old).
Her early works include a Ukrainian translation of Gogol’s “Evenings on a Farm” (co-authored with her brother). Ukrainka was published in foreign magazines “Zorya”, “Live and Slovo”, “Literary and Scientific Bulletin”.
Extraordinarily talented, receptive, vulnerable, with a deep, true musical talent (she began playing and composing small musical pieces from the age of five
!) at the age of eight, who wrote her first poem, Lesya unexpectedly fell seriously ill in 1881.
She was tormented by unbearable pain in her right leg. At first they decided that she had acute rheumatism, they treated her with baths, ointments, and herbs, but everything was useless. The pain went into my hands. Doctors were finally able to determine that it was bone tuberculosis
. Lesya's musical career was put to an end. After the first, complex, but extremely unsuccessful operation, my hand was left crippled! Then sadness first appeared in the eyes of the fragile girl. In the future, like a light blanket, it will envelop all her work. From now on, for many months of the year, the girl must lie in bed, not make sudden movements, and experience excruciating pain all the time...
Lesya Ukrainka always tried to find joy in everything, even in small things! She had an indomitable spirit. Selflessly, at night, she studied languages: Bulgarian, Spanish, Latin, ancient Greek, Italian, Polish, German, not to mention English and French, geography, the history of the East and oriental cultures, the history of art and religions, and for her younger sisters at the age of 19 wrote a textbook: “Ancient history of the eastern peoples.”
In 1893, in Lvov, (Western Ukraine), a thin book of her poems was published entitled: “On the Wings of Song” and was warmly received by critics and the public. The book quickly made her popular.
In 1898, in Zaporozhye, Larisa Petrovna met Sergei Konstantinovich Merzhinsky, a public figure, a graduate of the Kyiv University of St. Vladimir. Merzhinsky lived for some time in Yalta, undergoing treatment for tuberculosis. Four years later (in 1901), Lesya travels to winter Minsk to visit her terminally ill lover. In the difficult winter months, one of her strongest dramas, “Obsessed,” is born, Sergei Merzhinsky dies, and Larisa Petrovna forever puts on black mourning clothes.
Thirty-six years old, she fell in love again. A person who responded to her feelings with no less sincere and deep affection - Kliment Kvitka, a learned musicologist-folklorist, collector of folk legends and songs. Lesya’s mother was again vehemently against any relationship between her daughter “with some beggar,” as she contemptuously called Kliment - a man of gentle character, reserved, shy, who experienced a deep personal drama in childhood - he grew up in a foster family. But Kvitka became so passionately attached to a thin, sick woman with big sad eyes, who understood him perfectly, that he flatly refused to leave her! And, despite all the anger and gloomy forecasts for the future of the young people, Olga Petrovna was forced to agree to her daughter’s marriage. The young people made a decision: they refused the help of their parents. Clement earned all the money needed for the treatment of his seriously ill wife himself. They sold everything that could be sold: things, simple belongings, kitchen utensils. They only valued the library.
Lesya received treatment in Egypt and Greece, Germany and Austria. Everything was useless. Incurable kidney disease was added to the aggravated process of bone tuberculosis. She died in Surami (Georgia) on August 1, 1913
. She flew away “on the wings of song.” Her old dream came true: she always wanted to touch the clouds with her hands...
Lesya Ukrainka's literary activity took place during the era of imperialism and the first Russian revolution. In Ukrainian literature of that time, naturalistic, decadent-symbolist and revolutionary-democratic movements were identified. The latter, who was sharply opposed to all bourgeois-nationalist literature, was joined by P. Grabovsky, M. Kotsyubinsky and Teslenko. L. Ukrainian had a negative attitude towards both decadents and naturalists. Moving away from the liberals, she became more and more imbued with revolutionary ideas, approaching the revolutionary democrats.
The creative method of Lesya Ukrainka is closer to romanticism than to realism, but fundamentally it differed sharply from the method of the decadent Ukrainian neo-romantics, especially the symbolists, primarily in its ideological and thematic orientation, as well as artistic means. Unlike many decadent romantics, Lesya Ukrainka did not idealize the past, although she created her images on historical material; passionately hating the oppressive reality of Tsarist Russia, she did not indulge in pessimism, did not fall into despair, but, on the contrary, called for a struggle for complete liberation from all oppression and the destruction of the exploitation of man by man. Lesya Ukrainka's romanticism was permeated with revolutionary sentiment.
Works written by Lesya Ukrainka:
- a collection of poetry “On the Wings of Songs” (“On the Wings of Songs”, 1893);
- collection of poetry “Thoughts and Dreams” (“Dumi i mrii”, 1899);
- collection of poetry “Responses” (“Vіdguki”, 1902);
- Drama extravaganza “Forest Song” (“Forest Song”, 1911).
Information sources:
- Wikipedia: Lesya Ukrainka;
- Lesya Ukrainka: biographical information;
- Lesya Ukraine - biography;
- Lesya Ukrainka - short biography.
Visual analyzers
The afferent pathways of the central nervous system of the visual analyzer begin from the cones and rods of the retina of the eyeball. The impulses enter the optic chiasm as part of the optic nerves, and then along the tract are sent to the subcortical centers of the brain, that is, to the visual thalamus, geniculate lateral bodies and superior colliculus, located in the middle part of the brain.
In the midbrain, a response to these stimuli occurs, and in the nuclei of the thalamus an unconscious assessment of impulses begins that provide involuntary movements reproduced by a person. The main unconscious movements are running and walking. In the projection center of vision or in the calcarine sulcus of the occipital lobe of the brain, impulses arrive along the optic radiation from the geniculate bodies located in the internal capsule, after which a full analysis of the incoming data begins. In the cortex, which is adjacent to the calcarine sulcus, the central part responsible for visual memory, which is also called the associative visual center, finds its location.
Olfactory analyzer
The afferent pathway of the olfactory analyzer originates from the receptors of the mucous membrane, localized in the upper part of the nasal passage. After this, the impulses are sent to the axons of the olfactory bulbs, and they flow along the fibers of the olfactory nerves. Then the impulses are sent to the projection center of smell, which is located in the area of the parahippocampal gyrus and uncus. These impulses follow along the tract to the cortex of the temporal lobe of the brain. Most of the information received from the olfactory receptors is sent to the subcortical centers, which are located in the middle and intermediate parts of the brain. The subcortical centers of the brain, in response to olfactory stimuli, provide reflex regulation of muscle tone.
Based on this, it can be determined that the main feature of olfactory receptors is that nerve impulses initially enter the cortex of the cerebral hemispheres, and not the subcortical centers of smell. In this regard, a person first smells the smell, then begins to evaluate it, and only after that an unconscious coloring of the stimulus is formed in the brain on an emotional level. The entire process takes only a fraction of a second.
LECTURES ON NEUROPSYCHOLOGY: Lecture No. 4 5/5 (2)
Lecture 4. Violations of voluntary movements and actions.
- Motor analyzer: afferent and efferent mechanisms.
- Elementary movement disorders.
- The problem of apraxia.
Voluntary movements and actions are among the most complex mental functions of a person. Voluntary actions are a set of voluntary movements united by a single goal. Voluntary movements and actions are conditioned reflex in origin and are formed completely during life.
Modern psychological ideas about voluntary movements developed in the struggle both with the idealistic approach and with vulgar materialistic, mechanistic concepts (behaviorism, etc.). Important contributions to the modern understanding of voluntary movements were made by domestic physiologists (I.M. Sechenov, I.P. Pavlov, N.A. Bernshtein, P.K. Anokhin, etc.) and psychologists (L.S. Vygotsky, A.N. Leontiev, A. R. Luria,). Voluntary movements and actions can be both independent motor acts and means through which a variety of forms of behavior are realized.
Voluntary movements are part of oral and written speech, as well as many other higher mental functions. From a physiological point of view, voluntary movements include movements of the striated muscles of the arms, face of the legs, and the entire torso, i.e., the most extensive class of movements.
Modern physiology has a variety of information regarding the great complexity of both afferent and efferent mechanisms of voluntary movements. Afferent mechanism of voluntary movements afferentis - “bringing” Voluntary movements are complex afferent systems, including different types of afferentation, among which the basal one is kinesthetic afferentation.
Any movement is a complex multi-level system, where each level (or certain anatomical structures) is characterized by “leading afferentation” and its own set of regulated movements.
There are five levels of movement regulation: 1. rubro-spinal; 2. thalamo-pallidal; 3. pyramidal-striate; 4. parieto-premotor; 5. cortical “symbolic”.
All these levels combine involuntary and voluntary movements into a single system. The first and second levels are responsible for the regulation of involuntary movements (these include smooth muscle movements, tremor, tone, etc.).
The third to fifth levels are associated with the regulation of voluntary motor acts, which involve both movements of the entire body and individual parts of the body: arms, face, speech apparatus, etc.
Thus, voluntary movements are a whole set of different motor acts, regulated by different levels (structures) of the nervous system and controlled by different kinds of afferent impulses (and different “leading afferentations”).
Defeat of any of the listed levels leads to disturbances in movements at this level, as well as in those motor acts in which these movements are included as “background”. The type of afferentation, as well as the corresponding anatomical structures, are criteria for identifying a class of movements (this applies to both voluntary and involuntary movements). In other words, afferentation is the most important factor determining the type of movement.
A. R. Luria, analyzing observations of patients with local brain lesions, described the specific composition of the cortical zones involved in the brain organization of voluntary motor acts, including in the concept of “motor organizer” not only motor, but also sensory and associative cortical fields.
Luria noted that in addition to the actual motor zones of the cerebral cortex, many other cortical zones should be included in the cortical link of the motor analyzer, namely:
- postcentral parietal cortex, which provides analysis of cutaneous-kinesthetic afferentation coming from the organs of movement; 2. posterior occipital and parieto-occipital sections of the cerebral cortex, which provide regulation of movements using visual afferentation, and are also responsible for the spatial organization of movements; 3. temporal cortex (primarily the left hemisphere), which provides not only auditory-speech afferentation of speech motor skills, but also participates in all “verbalized” (external and internal speech) motor acts; 4. the anterior sections of the cerebral cortex (premotor and prefrontal cortex), with the help of which the programming of movements, the organization of movements in time and control over the implementation of the program are carried out.
Consequently, voluntary human movements are provided by a variety of types of afferentation, and therefore almost the entire cerebral cortex should be included in the cortical link of the motor analyzer. In general, modern neurophysiology and neuropsychology have developed the idea that voluntary movements are very complex afferent systems that are realized with the participation of almost the entire cerebral cortex.
Efferent mechanisms of voluntary movements (eff - efferent) The efferent (executive) mechanisms of voluntary movements and actions include, as is known, two interconnected, but to a certain extent autonomous efferent systems: pyramidal and extrapyramidal, the cortical sections of which form a single sensorimotor zone of the cortex.
The pyramidal system is the main mechanism that implements voluntary movements. It starts from the Betz motor cells located in the V layer of the motor cortex (4th field), continues in the form of the corticospinal, or pyramidal, tract, which passes to the opposite side in the area of the pyramids and ends on the motor neurons of the spinal cord (2nd field). pyramidal tract neuron) innervating the corresponding muscle group. New data have been added to these ideas about the pyramidal system as the main efferent mechanism of voluntary movements:
Firstly, not only the 4th field is motor. Pyramidal-type motor cells were found in fields 6 and 8 of the precentral cortex, and in fields 2, 1 and 3 of the postcentral cortex. Consequently, the pyramidal path begins not only from the 4th field, as previously assumed, but from significantly larger areas of the cerebral cortex. Only 40% of all fibers of the pyramidal tract begin in the 4th area, about 20% in the postcentral gyrus; the rest are in the premotor zone of the cerebral cortex.
Secondly, it has been established that the pyramidal tract contains fibers of various types (in diameter and degree of myelination). Well myelinated fibers make up no more than 10% of all pyramidal fibers that extend from the cortex to the periphery.
Thirdly, if it was previously assumed that there is a single pyramidal, or corticospinal, tract (lateral), which crosses in the pyramidal zone from the cerebral cortex to the motor neurons of the spinal cord, now another corticospinal tract has been identified ( ventral), running without decussation as part of the pyramids on the same side. Fourthly, the pyramidal tract does not end directly on motor neurons located in the anterior horns of the spinal cord, as previously thought, but mainly on intermediate (or intercalary) neurons, with the help of which the excitability of the main motor neurons is modulated and thereby affects the final result - voluntary movements. All these data indicate the complexity of the pyramidal system as an actuator.
In addition to the motor zones of the cerebral cortex, the irritation of which causes movements, there are also zones of the cortex, the irritation of which stops movement that has already begun (suppressive areas of the cortex).
In the cerebral cortex there are also special aggressive zones. These are areas of the cortex that are well known to neurosurgeons and neurologists. Their irritation (by electric current or a painful process) causes adversive epileptic seizures (starting with adversia - turning the body, eyes, head, arms and legs in the direction opposite to the location of the exciting agent). Epilepsy occurring with this type of seizure is known as Jacksonian epilepsy.
There are two aggressive cortical zones: premotor and parieto-occipital. (fields 6, 8 and 19). These cortical fields are involved in reactions associated with attention to a stimulus, that is, in the organization of complex motor acts that mediate attention to a specific stimulus.
The extrapyramidal system is the second efferent mechanism for the implementation of voluntary movements and actions. The extrapyramidal system refers to all motor pathways that do not pass through the pyramids of the medulla oblongata.
The extrapyramidal system is more ancient than the pyramidal system. It is very complex in its composition. In the extrapyramidal system, cortical and subcortical sections are distinguished. The cortical section of the extrapyramidal system includes the same fields that are included in the cortical nucleus of the motor analyzer. These are the 6th and 8th, as well as the 1st and 2nd fields, i.e., the sensorimotor area of the cortex.
The structure of the subcortical section of the extrapyramidal system is quite complex. It consists of a number of formations. The final authority of extrapyramidal influences are the same motor neurons of the spinal cord to which impulses from the pyramidal system are addressed.
!!!! There is no clear anatomical boundary between the pyramidal and extrapyramidal systems. They are isolated anatomically only in the area of the pyramids, in the medulla oblongata. However, the functional differences between these systems are quite distinct. And they manifest themselves especially clearly in the clinic of local brain lesions.
Disorders of motor functions that arise from various local brain lesions can be divided into relatively elementary ones, associated with damage to the executive, efferent mechanisms of movements, and more complex ones, extending to voluntary movements and actions and associated mainly with damage to the afferent mechanisms of motor acts.
Relatively elementary movement disorders occur when the subcortical parts of the pyramidal and extrapyramidal systems are damaged. When the cortical part of the pyramidal system (4th field), located in the precentral region, is damaged, movement disorders are observed in the form of paresis or paralysis of a certain muscle group: the arm, leg or torso on the side opposite to the lesion. Lesion of the 4th field is characterized by flaccid paralysis (when the muscles do not resist passive movement), occurring against the background of decreased muscle tone. But with foci located anterior to the 4th field (in the 6th and 8th fields of the cortex), a picture of spastic paralysis appears, i.e., loss of corresponding movements against the background of increased muscle tone. The phenomena of paresis, together with sensory disorders, are also characteristic of damage to the post-central parts of the cortex. These motor dysfunctions are studied in detail by neurology. Along with these neurological symptoms, damage to the cortical part of the extrapyramidal system also causes disturbances in complex voluntary movements, which will be discussed below.
When the pyramidal tracts are damaged in the subcortical areas of the brain (for example, in the area of the internal capsule), complete loss of movements (paralysis) occurs on the opposite side. Complete unilateral loss of movements of the arms and legs (hemiplegia) appears with rough lesions. More often, in the clinic of local brain lesions, phenomena of partial decrease in motor functions on one side (hemiparesis) are observed.
When crossing the pyramidal tract in the pyramidal zone - the only zone where the pyramidal and extrapyramidal tracts are anatomically separated - voluntary movements are realized only with the help of the extrapyramidal system. When crossing pyramids, movements become less discrete (i.e., more coarse). More muscles begin to participate in grasping objects. In addition, in movements that require the participation of all muscles (such as jumping), which is typical for the motor skills of monkeys (surgeries were performed on them), accuracy and coordination disappear. Such movement disorders occur against the background of increased muscle tone. These data suggest that the pyramidal system is involved in the organization of predominantly precise, discrete, spatially oriented movements and in the suppression of muscle tone. Damage to the cortical and subcortical parts of the extrapyramidal system leads to the appearance of various movement disorders. These disorders can be divided into dynamic (i.e., disturbances in actual movements) and static (i.e., disturbances in posture).
When the cortical level of the extrapyramidal system is damaged (fields 6 and 8 of the premotor cortex), spastic motor disorders occur in the contralateral limbs. Stimulation of the 6th or 8th fields causes turns of the head, eyes and body in the opposite direction (adversia), as well as complex movements of the contralateral arm or leg.
Damage to the subcortical striopallidal system, caused by various diseases (parkinsonism, Alzheimer's disease, Pick's disease, tumors, hemorrhages in the area of the basal ganglia, etc.), is characterized by general immobility, adynamia, and difficulty moving. At the same time, violent movements of the contralateral arms, legs, and head appear—hyperkinesis. In such patients, there is a violation of tone (in the form of spasticity, rigidity or hypotonia), which forms the basis of the posture, and a violation of motor acts (in the form of increased tremor - hyperkinesis). Patients lose the ability to care for themselves and become disabled. Currently, surgical (stereotactic) and conservative (pharmacological) methods for treating parkinsonism have been developed. The essence of the first is the destruction of one of the “sick” extrapyramidal structures (for example, the ventrolateral nucleus of the thalamus) and the cessation of pathological excitation circulating along closed extrapyramidal pathways. Depending on the form of parkinsonism, either the pallidum zone or the striatum zone (in the area of the caudate nucleus or putamen) is also destroyed. The pallidum zone (globus pallidus) is related to the regulation of slow smooth movements and the maintenance of tone (posture), while the striatum zone (putamen and caudate nucleus) is more associated with physiological tremor, which in pathology turns into hyperkinesis. Selective damage to the pallidum zone (an older part than the striatum) can lead to athetosis or choreoathetosis (pathological wave-like movements of the arms and legs, twitching of the limbs, etc.).
The defeat of striopallidal formations is accompanied by another type of motor symptoms - a violation of facial expressions and pantomimics, i.e., involuntary motor components of emotions. These disturbances can appear either in the form of amymia (mask-like face) and general immobility, or in the form of forced laughter, crying or forced walking, running (propulsion). Often these patients suffer from subjective experience of emotions. Finally, in such patients, physiological synergies are also disrupted - normal combined movements of different motor organs (for example, swinging arms while walking), which leads to unnaturalness of their motor acts.
The consequences of damage to other structures of the extrapyramidal system have been studied to a lesser extent, with the exception, of course, of the cerebellum. The cerebellum is the most important center for coordinating various motor acts, an “organ of balance” that provides a number of unconditional motor acts associated with visual, auditory, cutaneous-kinesthetic, and vestibular afferentation. Damage to the cerebellum is accompanied by a variety of movement disorders (primarily disorders of coordination of motor acts). Their description constitutes one of the well-developed sections of modern neurology.
Damage to the pyramidal and extrapyramidal structures of the spinal cord is reduced to a dysfunction of motor neurons, as a result of which the movements controlled by them are lost (or disrupted). Depending on the level of damage to the spinal cord, the motor functions of the upper or lower extremities are impaired (on one or both sides), and all local motor reflexes are carried out, as a rule, normally or are even enhanced due to the elimination of cortical control. All these movement disorders are also discussed in detail in the neurology course.
Clinical observations of patients who have damage to one or another level of the pyramidal or extrapyramidal system have made it possible to clarify the functions of these systems. The pyramidal system is responsible for the regulation of discrete, precise movements, completely subordinate to voluntary control and well afferented by “external” afferentation (visual, auditory). It controls complex spatially organized movements in which the whole body is involved. The pyramidal system regulates predominantly the phasic type of movements, i.e. movements that are precisely dosed in time and space.
The extrapyramidal system controls mainly the involuntary components of voluntary movements; In addition to the regulation of tone (the background of motor activity against which phasic short-term motor acts are played out), these include: • maintaining posture; • regulation of physiological tremor; • physiological synergies; • coordination of movements; • general coordination of motor acts; • their integration; • body plasticity; • pantomime; • facial expressions, etc.
The extrapyramidal system also controls a variety of motor skills and automatisms. In general, the extrapyramidal system is less corticolized than the pyramidal system, and the motor acts regulated by it are less voluntary than the movements regulated by the pyramidal system. It should, however, be remembered that the pyramidal and extrapyramidal systems represent a single efferent mechanism, the different levels of which reflect different stages of evolution. The pyramidal system, as an evolutionarily younger system, is to a certain extent a “superstructure” over the more ancient extrapyramidal structures, and its emergence in humans is primarily due to the development of voluntary movements and actions. Disorders of voluntary movements and actions
Disturbances of voluntary movements and actions are complex movement disorders, which are primarily associated with damage to the cortical level of motor functional systems. This type of motor dysfunction is called apraxia in neurology and neuropsychology. Apraxia refers to such disorders of voluntary movements and actions that are not accompanied by clear elementary movement disorders - paralysis and paresis, obvious disorders of muscle tone and tremor, although combinations of complex and elementary movement disorders are possible.
Apraxia primarily refers to disorders of voluntary movements and actions performed with objects. The difficulties of understanding the nature of apraxia are reflected in their classifications. The most famous classification, proposed at one time by G. Lipmann, distinguishes three forms of apraxia: • ideational, suggesting the disintegration of the “idea” of movement, its concept; • kinetic, associated with a violation of the kinetic “images” of movement; • ideomotor, which is based on the difficulties of transferring “ideas” about movement to “movement execution centers.” G. Lipmann associated the first type of apraxia with diffuse brain damage, the second with damage to the cortex in the lower premotor region, and the third with damage to the cortex in the lower parietal region.
A. R. Luria developed a classification of apraxia based on a general understanding of the psychological structure and brain organization of a voluntary motor act. He identified four forms of apraxia
The first is kinesthetic apraxia. This form of apraxia occurs when the lower parts of the postcentral region of the cerebral cortex are damaged. In these cases, there are no clear motor defects, muscle strength is sufficient, but the kinesthetic basis of movements suffers. They become undifferentiated and poorly controlled (the “shovel hand” symptom). Patients have impaired movements when writing, the ability to correctly reproduce various hand postures (postural apraxia); They cannot show without an object how this or that action is performed (for example, how tea is poured into a glass, how a cigarette is lit, etc.). While the external spatial organization of movements is preserved, the internal proprioceptive kinesthetic afferentation of the motor act is disrupted. With increased visual control, movements can be compensated to a certain extent. When the left hemisphere is damaged, kinesthetic apraxia is usually bilateral in nature; when the right hemisphere is damaged, it often manifests itself only in one left hand.
The second form of apraxia, spatial apraxia, or apraktoagnosia, occurs with damage to the parieto-occipital cortex at the border of the 19th and 39th fields. The basis of this form of apraxia is a disorder of visual-spatial synthesis, a violation of spatial representations (“top-bottom”, “right-left”, etc.). Thus, in these cases, visuospatial afferentation of movements is affected. Spatial apraxia is more often observed in combination with visual opto-spatial agnosia. Then a complex picture of apraktoagnosia arises. In all cases, patients experience apraxia of posture and difficulties in performing spatially oriented movements (for example, patients cannot make the bed, get dressed, etc.). Strengthening visual control of movements does not help them.
The third form of apraxia - kinetic apraxia - is associated with damage to the lower parts of the premotor area of the cerebral cortex (the 6th, 8th fields - the anterior sections of the “cortical” nucleus of the motor analyzer). It occurs against the background of a violation of automation (temporal organization) of various mental functions. Manifests itself in the form of the disintegration of “kinetic melodies”, i.e. a violation of the sequence of movements, the temporary organization of motor acts. This form of apraxia is characterized by motor perseverations, manifested in the uncontrolled continuation of a movement that has once begun.
Kinetic apraxia manifests itself in a violation of a wide variety of motor acts: object actions, drawing, writing, and in the difficulty of performing graphic tests, especially with the serial organization of movements (dynamic apraxia). With damage to the lower premotor cortex of the left hemisphere (in right-handed people), kinetic apraxia is observed, as a rule, in both hands.
The fourth form of apraxia—regulatory or prefrontal apraxia—occurs when the convexital prefrontal cortex is damaged anterior to the premotor regions; occurs against the background of almost complete preservation of tone and muscle strength.
It manifests itself in the form of violations of the programming of movements, the disabling of conscious control over their execution, and the replacement of necessary movements with motor patterns and stereotypes. With a gross breakdown of voluntary regulation of movements, patients experience symptoms of echopraxia in the form of uncontrolled imitative repetitions of the experimenter’s movements.
Regulatory apraxia is characterized by systemic perseverations, i.e., perseverations of the entire motor program as a whole, and not of its individual elements. Such patients, after writing under dictation in response to a proposal to draw a triangle, trace the outline of the triangle with movements characteristic of writing, etc. The greatest difficulties in these patients are caused by changing programs of movements and actions. The basis of this defect is a violation of voluntary control over the implementation of movement, a violation of speech regulation of motor acts.
The classification of apraxia created by A. R. Luria is based mainly on the analysis of motor dysfunction in patients with damage to the left hemisphere of the brain. The forms of disturbance of voluntary movements and actions with damage to various cortical zones of the right hemisphere have been studied to a lesser extent; This is one of the urgent tasks of modern neuropsychology.
Vestibular tract
The vestibular afferent pathway begins from the receptors of the semicircular canal of the inner ear, the uterus and the receptors that make up this organ. This tract in the central nervous system is responsible for coordinating movements and maintaining balance during physical and vestibular stress.
Afferent centripetal pathways and the peculiarity of their structure indicate that a person needs to make a lot of effort to maintain the health and integrity of each organ individually and together. Each component of this pathway provides the body with all the necessary information, helps to immediately process it and carry out all vital processes. This is important in the functioning of the entire organism as a whole and individual organs.