In the medulla oblongata, the axons of second neurons pass between the inferior olive and the inferior cerebellar peduncle. In the tegmentum of the bridge they approach its dorsal surface. At the border with the midbrain, the fibers of the anterior spinocerebellar tract turn dorsally into the region of the superior medullary velum. Here, part of the fibers once again passes to the opposite side, and then through the superior cerebellar peduncles (pedunculi cerebellares superiores) they reach the anterosuperior parts of the cerebellum and enter the nucleus of the tent, nucleus fastigii, cerebellar vermis, vermis cerebelli. ). The tent nucleus is a collector of afferent information. From here it goes to the piriform neurons of the cerebellar cortex - Purkinje cells. Jan Evangelista Purkinje, 1787-1869, Czech physiologist, histologist, embryologist and pharmacologist. Pear-shaped neurons process the received information and transmit it to the dentate nucleus (nucleus dentatus) of the cerebellar hemisphere of the same name. The afferent pathways end in this structure. The dentate nucleus is the final link of the cerebellar regulators. Its neurons form and send efferent (control) signals through the superior cerebellar peduncles to the red nucleus of the midbrain on the opposite side (Wernecking's chiasm). It is possible to trace the fiber systems through which information from the vermis cortex reaches the red nucleus, the cerebellar hemisphere, as well as the overlying parts of the brain - the cerebral cortex. From the cortex of the vermis, through the corky and globular nuclei, information through the superior cerebellar peduncle is sent to the red nucleus of the opposite side (cerebellotegmental tract). The vermis cortex is connected by associative fibers to the cerebellar hemisphere cortex, from where information enters the dentate nucleus of the cerebellum. There are also connections between the cerebellum and the cortex through the thalamus. In particular, the axons of the neurons of the dentate nucleus exit through the superior cerebellar peduncle into the tegmentum of the bridge, pass to the opposite side and go to the thalamus. Having switched to the next neuron in the thalamus, the paths follow to the cerebral cortex, to the postcentral gyrus. The spinocerebellar tracts described are the main ones, but not the only known spinocerebellar tracts. The sphenocerebellar tract (tractus cuneocerebellaris) is considered part of the posterior spinocerebellar tract. The rostral spinocerebellar tract (tractus spinocerebellaris rostralis) is considered part of the anterior spinocerebellar tract. There are also indirect spinocerebellar tracts. Among them, the most studied are: spino-olivary-cerebellar tracts, spino-bulbo-cerebellar tracts and spino-reticulo-cerebellar tracts.
Thanks to information arriving along the anterior spinal cerebellar tracts and posterior spinal cerebellar tracts, the cerebellum, as one of the nerve centers in the hierarchy of regulators of the musculoskeletal system, is involved in controlling movements that ensure body balance without the participation of higher parts of the brain (cerebral cortex) brain), without the participation of consciousness.
See: Neurology: Dictionary, Neurology: Internet Resources.
| LIBRARY = LIBRARY Human Physiology = Human Physiology, Human Anatomy = Human Anatomy, Human Biochemistry = Human Biochemistry, Human Psychology = Human Psychology, Medicine = Medicine, Mathematics = Mathematics, Chemistry = Chemistry, Physics = Physics, General Scientific Literature = General Science Lexis. Click here to access any source in the site's library! Click here and receive access to the any reference of the library! “I AM LEARNED. . . N E D O U C H A ?” T E S T V A S H E G O I N T E L L E C T A Premise : The effectiveness of the development of any branch of knowledge is determined by the degree of compliance with the methodology of knowledge - the knowable entity. |
Error? Click here and fix it! Search on the site E-mail of the author (author)
Anatomy of the nervous system pathways | Publishing house PIMU – Part 3
Three of them are localized in the ampoules of the semicircular ducts and are called ampullary ridges, another two are located in the uterus and sac and are called spots.
Both ampullary ridges and spots are formed by a cluster of special sensory hair cells of an epithelial nature, participating in the lining of the ampullae, spherical and elliptical sacs and associated with sensory nerve endings.
The conductor of the vestibular analyzer is a chain of three neurons. The body of the first neuron is located in the vestibular node, located deep in the internal auditory canal and formed by bipolar neurons.
Both peripheral and central processes of these ganglion cells participate in the formation of the vestibular part (vestibular nerve) of the vestibulocochlear nerve.
The peripheral processes form synaptic contact with receptor hair cells, and the central processes in the pontocerebellar angle (behind the middle cerebellar peduncles) “enter” the substance of the pons, are divided into ascending and descending and “approach” the vestibular nuclei (ascending - to the superior nucleus, and descending ones - to the lateral, medial and inferior nuclei). The body of the second neuron is localized in the lateral and medial vestibular nuclei. The axons of the second neurons at the level of the bridge make a “transition” to the opposite side and end in the median nuclei of the thalamus, where the bodies of the third neurons are located. Their axons form a thalamocortical tract, “passing” through the middle of the posterior limb of the internal capsule and ending in the cerebral cortex.
The cortical end of the vestibular analyzer, according to most authors, is located in the cortex of the parietal and (or) temporal lobes. At the same time, versions are expressed in favor of the superior parietal lobule, supramarginal and postcentral gyri, as well as the superior, middle and inferior temporal gyri.
Diseases and lesions of the organ of balance and the vestibular nuclei of the pons lead to imbalance, deviation of the body to the side, dizziness, rhythmic twitching of the eyeballs (nystagmus) and autonomic disorders (nausea, vomiting, constriction of the pupils, decreased pulse, decreased blood pressure and cold extremities, increased sweating, increased intestinal motility). The clinical picture of damage to the cortical end of the vestibular analyzer is complemented by a violation of orientation in space.
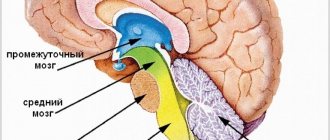
2.2. Proprioceptive pathways to the cerebellum
These pathways relate to the pathways of unconscious proprioceptive sensitivity, which may have served as the main factor in the formation of such an ancient center of balance in the evolutionary sense, which is the cerebellum. Within the central nervous system, direct projections from the spinal cord to the cerebellum and those mediated through the reticular formation and the olive nucleus of the medulla oblongata are distinguished.
The most important are the direct projections, called the “anterior and posterior spinocerebellar tracts.”
They transmit proprioceptive information from skeletal muscles, bones, joint capsules and ligaments, allowing the cerebellum to assess the state of the musculoskeletal system of the torso, neck and limbs and regulate muscle tone and coordinate movements aimed, in particular, at maintaining balance.
The posterior spinocerebellar tract is more differentiated in a functional sense, as it carries information from each individual muscle. The anterior spinocerebellar tract transmits more general information about the state of a particular group of synergistic muscles. The spinocerebellar tracts originate in skeletal muscles, bones, and bone joints with proprioceptors.
Posterior spinocerebellar tract (bundle of Flexig)
The dorsal (posterior thoracic) nucleus, localized at the base of the dorsal horn, is clearly expressed in the spinal segments C7–L2. Within the spinal cord tr. spinocerebellaris posterior, receiving fibers segment by segment, thickens to the level of C7. Both in the lateral funiculus of the spinal cord and in the inferior cerebellar peduncle (Fig. 5), the Flexig bundle occupies a superficial dorsolateral position.
Rice. 5. Posterior spinocerebellar tract: 1 - muscle, 2 - spinal ganglia (I), 3 - dorsal (posterior thoracic) nucleus of the spinal cord (II), 4 - posterior spinocerebellar tract, 5 - inferior cerebellar peduncle, 6 - cortex of the lower part of the vermis (III)
The path ends in the lower part of the worm, in its bark, both on its own and, according to some authors, on the opposite side.
Anterior spinocerebellar tract (Gowers bundle)
The difference between this pathway is the localization of the body of the second neuron in the nucl. intermediomedialis, located in the central intermediate substance of the spinal cord. Most (90%) of the neurons in this nucleus send their axons through the commissure alba to the opposite side (Fig. 6).
Rice. 6. Anterior spinocerebellar tract: 1 - muscle group, 2 - spinal nodes (I), 3 - intermedial nucleus of the spinal cord (II), 4 - white commissure, 5 - anterior spinocerebellar tract, 6 - superior medullary velum, 7 - superior cerebellar peduncle, 8 - cortex of the superior part of the vermis (III)
A smaller part (10%) is sent to the Govers beam of its side. Thus, in every tr. spinocerebellaris anterior there are fibers from its own and the opposite side, and the latter predominate quantitatively.
//www.youtube.com/watch?v=D4ii1lc9PzE
The Govers' bundle in the spinal cord is located superficially in the ventrolateral part of the lateral funiculus, with fibers from the overlying segments added on the lateral side. In the medulla oblongata tr. spinocerebellaris anterior occupies a place between the olive and the inferior cerebellar peduncle, and then “passes” into the lateral part of the pontine tegmentum.
Near the rostral border of the pons, the Gowers bundle “shifts” dorsally and through the superior medullary velum, the fibers that crossed in the spinal cord “return” to their side and through the superior cerebellar peduncle reach their end point in the cortex of the upper part of the vermis.
Thus, unconscious proprioceptive sensitivity is transmitted to the cerebellum on the same side of the body.
Common to the described proprioceptive pathways is the localization of the body of the first neuron in the spinal ganglion, the second in the nuclei of the spinal cord, and the third in the cortex of the cerebellar vermis.
2.3. Pathway of conscious proprioceptive sensitivity (“motor” analyzer)
The proprioceptive field (a set of proprioceptors), the path of conscious proprioceptive sensitivity and the area of the cerebral cortex into which the receptive field of the musculoskeletal system is projected were previously united under the name “motor analyzer”. This name is clearly unfortunate, since it contradicts the very idea of I.P.
Pavlov, who described the analyzer as an exclusively sensory, sensitive system. Therefore, if we use the term “motor analyzer,” it should be interpreted as an analyzer whose receptor field is located in the structures of the musculoskeletal system, including skeletal muscles.
It would be more correct to call this analyzer proprioceptive, but so far this term has not become widespread.
The “motor” (proprioceptive) analyzer evaluates muscle tone, the position of body parts in space, the presence of gravity and vibration.
Receptors are localized in skeletal muscles, bones, and bone joints.
The conductor is a chain of three neurons. The body of the first pseudounipolar neuron is found in all spinal ganglia. Peripheral processes participate in the formation of spinal nerves and their branches that innervate the organs of the musculoskeletal system (Fig. 7).
Rice. 7. “Motor” (proprioceptive) analyzer: 1 - muscles of the neck, upper torso and upper limbs, 2 - muscles of the lower torso and lower extremities, 3 - spinal nodes (I), 4 - thin fasciculus of Gaulle, 5 - wedge-shaped fasciculus Burdakha, 6 - thin nucleus (II), 7 - cuneate nucleus (II), 8 - internal arcuate fibers, 9 - medial lemniscus, 10 - ventrolateral nuclei of the thalamus (III), 11 - thalamocortical pathway, 12 - precentral gyrus, postcentral gyrus , superior parietal lobule (IV)
Source: //medread.ru/anatomiya_provodyashhix_putej_nervnoj_sistemy/3/
Posterior and anterior spinocerebellar tracts
The previous article examined the path of nerve impulses from the peripheral parts of the nervous system
to the spinal cord, and now consider the further passage of nerve impulses through the central nervous system.
Entrance zone of nerve roots and dorsal horn
. Sensory fibers enter the spinal cord through the dorsal root input zone (Redlich-Obersteiner area) and then send off numerous collaterals that form synaptic contacts with other neurons in the spinal cord. Each bundle of fibers belonging to a specific type of sensitivity is assigned its own place in the spinal cord.
It should be noted that as the afferent fibers pass
through the entrance zone of the root and the dorsal horn, their myelin sheath becomes significantly thinner.
Moreover, the type of myelin also changes from the peripheral parts of the nervous system to the central ones, since there myelin is no longer formed by lemmocytes, but by oligodendrocytes.
Now let's look in detail at each of the afferent (ascending) pathways
, carrying impulses of various types of sensitivity in the spinal cord.
Some nerve impulses
, originating in the musculoskeletal system (muscles, tendons and joints), ascend along the spinocerebellar tracts to the cerebellum, which is the center of balance and coordination. Each half of the spinal cord contains two spinocerebellar tracts - anterior and posterior.
Posterior spinocerebellar tract (flexig tract)
. After fast-conducting group IA fibers from the muscle spindles and tendon organs enter the spinal cord, they divide into numerous collaterals. Some of these collateral fibers form direct synaptic contacts with the bodies of large a-motoneurons in the anterior horns (monosynaptic reflex arc).
Other collateral fibers
, ascending from the thoracic, lumbar and sacral levels, ending in a pillar-like nucleus lying at the base of the dorsal horns between segments C8-L2 and known variously as the lateral intermediate column, thoracic nucleus, thoracic column, Clark's column or Stilling's nucleus.
Second (postsynaptic) neurons
, the bodies of which are contained in this nucleus, give rise to the posterior spinocerebellar tract, the fibers of which belong to one of the most rapidly conducting types.
The posterior spinocerebellar tract ascends on its (ipsilateral) side of the spinal cord along the posterior part of the lateral funiculus and then, as part of the inferior cerebellar peduncle, enters the cerebellar vermis.
Afferent fibers that enter the spinal cord at the level of the cervical region, that is, above the lateral intermediate column, follow as part of the cuneate fasciculus and form synaptic contacts on second neurons in the accessory cuneate nucleus of the medulla oblongata, from where their axons extend into the cerebellum.
Anterior spinocerebellar tract (Gowers tract)
. Other group IA afferents entering the spinal cord synapse with funicular neurons in the dorsal horn and central gray matter of the spinal cord.
The mentioned second neurons, which are already found in the lower lumbar segments, give rise to the anterior spinocerebellar tract, which ascends both on its own and on the opposite side of the spinal cord and ends in the cerebellum.
Anterior spinocerebellar tract
, rising to the midbrain, passes under the bottom of the IV ventricle, turns posteriorly above it and, unlike the posterior spinocerebellar tract, penetrates the cerebellar vermis through the superior cerebellar peduncle and the superior medullary velum.
The cerebellum receives afferent impulses of proprioceptive sensitivity from all areas of the body; its polysynaptic efferent fibers, in turn, influence muscle tone and coordination of agonist and antagonist muscles (synergistic muscle activity), which are involved in maintaining an upright body position during standing and walking, as well as performing other movements.
So the movements
are regulated by various feedback systems, some of which close at the level of the spinal cord, others at higher levels of the central nervous system and involve extrapyramidal pathways. Feedback systems affecting higher levels of the central nervous system involve both α and γ motor neurons. All these processes occur without the participation of consciousness.
- Read "
Posterior funiculi of the spinal cord. Signs of damage to the posterior cords”
Contents of the topic “Neurology”:
Source: //dommedika.com/nevrologia/spinnomozgechkovie_puti.html