Центростремительные волокна
Афферентные нервные пути классифицируются на пути бессознательной и сознательной чувствительности. Именно при помощи них обеспечивается связь между всеми интеграционными центрами, расположенными в головном мозге. К примеру, они обеспечивают прямую связь между мозжечком и корой полушарий большого мозга.
Основные афферентные пути ЦНС сознательной общей чувствительности – это волокна болевой, температурной и тактильной чувствительности, а также сознательной проприоцептивной. Основные бессознательные проводящие пути общей чувствительности – это передний и задний спинно-мозжечковые. К специальным проводящим относятся вестибулярный, слуховой, вкусовой, обонятельный и зрительный.
ОРГАНЫ ЧУВСТВ
Некоторые закономерности строения эфферентных проекционных проводящих путей
1. Первый нейрон всех эфферентных путей локализуется в коре большого мозга.
2. Эфферентные проекционные пути занимают переднюю ножку, колено и переднюю часть задней ножки внутренней капсулы, проходят в основании ножек мозга и моста.
3. Все эфферентные пути заканчиваются в ядрах двигательных черепных нервов и в передних рогах спинного мозга, где располагается последний, двигательный нейрон.
4. Эфферентные пути образуют полный или частичный перекрест, вследствие чего импульсы из коры полушария большого мозга передаются мышцам противоположной половины тела.
Органы чувств осуществляют восприятие различных раздражений, действующих на организм человека и животных, а также первичный анализ этих раздражений. Академик И.П.Павлов определял органы чувств как периферические зоны анализаторов. Их специфическими воспринимающими элементами являются чувствительные нервные окончания — рецепторы, преобразующие энергию внешнего раздражителя в нервные импульсы. Последние содержат в закодированной форме информацию о предметах и явлениях внешнего мира. Эти импульсы передаются по афферентным нервным путям подкорковым и корковым центрам, где происходит окончательный анализ раздражений. Согласно учению об анализаторах, афферентные пути представляют собой их средний, проводниковый отдел, а воспринимающие зоны коры являются их центральными концами. С корковыми отделами анализаторов связано возникновение ощущений.
У простейших чувствительность присуща наружному слою протоплазмы их единственной клетки. У низших животных, тело которых состоит из энтодермы и эктодермы, все клетки последней отвечают на внешние раздражения. Одновременно с дифференцировкой мышечной и нервной системы в эктодерме обособляются отдельные перцептирующие клетки, которые связаны с центральной нервной системой и представляют собой первичные чувствующие клетки: они вначале (у низших кишечнополостных) рассеяны по всему телу, затем группируются в определенных местах, особенно в окружности рта. Такие группы чувствующих клеток — простейшие по устройству и функциям органы чувств. Наконец, более совершенные формы наблюдаются у высших, где в состав органов чувств входят не только перцептирующие элементы, но и особые добавочные (вспомогательные) аппараты: сначала — индифферентные (опорные) эпителиальные клетки, затем соединительная и мышечная ткани.
В процессе эволюции развиваются органы, приспособленные для восприятия самых разнообразных агентов окружающей среды — механических, физических, химических. Например, термиты воспринимают магнитное поле, пчелы и муравьи — ультрафиолетовые лучи, тараканы и кальмары — инфракрасные лучи, рыбы имеют орган боковой линии, воспринимающий направление и скорость движения воды, а землеройки и летучие мыши способны к восприятию ультразвуковых колебаний. У высших животных и человека органами чувств являются орган обоняния, орган вкуса, орган зрения, преддверно-улитковый орган и кожа, которая вместе с ее придатками образует общий покров тела.
На основании особенностей развития, строения и функции выделяют 3 типа органов чувств. К I типу относятся органы зрения и обоняния, которые закладываются у эмбриона как часть мозга. В основе их строения лежат первично-чувствующие, или нейросенсорные, клетки. Эти клетки имеют специализированные периферические отростки, воспринимающие колебания световых волн или молекул летучих веществ, и центральные отростки, по которым возбуждение передается афферентным нейронам.
Ко II типу принадлежат орган вкуса, слуха и равновесия. Они закладываются в эмбриональном периоде в виде утолщения эктодермы, плакод. Их основным рецепторным элементом являются вторично-чувствующие, сенсоэпителиальные клетки. В отличие от нейросенсорных клеток они не имеют аксоноподобных отростков. Возбуждение, возникающее в них под действием вкусовых веществ, колебаний воздуха или жидкой среды, передается окончаниям соответствующих нервов.
Третий тип органов чувств представлен рецепторными инкапсулированными или неинкапсулированными тельцами и образованиями. К ним относятся рецепторы кожи и подкожной ткани. Они представляют собой нервные окончания, окруженные соединительнотканными или глиальными клетками. Общим признаком всех воспринимающих клеток является наличие у них жгутиков — киноцилий или микроворсинок — стереоцилий. В плазматическую мембрану жгутиков и микроворсинок вмонтированы молекулы особых фото-, хемо- и механорецепторных белков. Эти молекулы воспринимают воздействия лишь одного определенного вида и кодируют их в специфическую информацию клетки, которая передается соответствующим нервным центрам.
Органы чувств различаются также по сложности своего анатомического строения. Относительно просто устроены органы вкуса и кожного чувства, которые представлены в основном эпителиальными образованиями. Органы обоняния, зрения, слуха и равновесия имеют вспомогательные аппараты, обеспечивающие поступление к ним только тех раздражений, к восприятию которых приспособлены эти органы чувств. Так вспомогательным аппаратом органа обоняния является решетчатый лабиринт и околоносовые пазухи, направляющие воздушную струю к обонятельным рецепторам. Орган зрения снабжен оптическим аппаратом, отбрасывающим изображение внешних предметов на сетчатку глаза. Орган слуха обладает сложно устроенным аппаратом улавливания и проведения звуков.
Вспомогательные аппараты органов чувств не только обеспечивают взаимодействие специфических раздражителей с рецепторами, но и преграждают путь посторонним, неадекватным раздражениям, а также обеспечивают защиту органов чувств от внешних механических воздействий и повреждений.
Волокна тактильной, температурной и болевой чувствительности
Данный путь берет свое начало с рецепторов в эпителии, импульсы от которых поступают в клетки спинномозгового узла, а после этого в спинной мозг, к ядрам зрительного бугра. Затем к коре постцентральной извилины, в которой и происходит их полный анализ. В данный проводящий путь вовлечено три тракта:
- Таламо-корковый.
- Ганглиоспинномозговой.
- Боковой спинно-таламический тракт, который проходит в боковом канатике спинного мозга и покрышке ствола головного мозга.
За получение тактильных ощущений в лицевой части головы и изменений температуры тела отвечает тройничный нерв. При его повреждении у человека начинаются сильные боли в области лица, которые то исчезают, то вновь появляются. Тройничный нерв проходит через шейный отдел, там, где перекрещиваются моторные волокна кортикоспинального тракта. Аксоны чувствительных нейронов тройничного нерва проходят через одну из частей продолговатого мозга. Через эти аксоны в мозг поступает информация о болевых ощущениях в ротовой полости, зубах, а также в верхней и нижней челюстях.
Проекционные и ассоциативные зоны коры
text_fields
text_fields
arrow_upward
Тела афферентных нейронов, как правило, лежат в чувствительных спинно-мозговых или черепно-мозговых ганглиях. Исключение составляют зрительная и обонятельная системы, где чувствительные нейроны находятся непосредственно в сетчатке (ганглиозные клетки) или обонятельной луковице соответственно. Отростки этих нейронов входят в спинной или головной мозг, где происходит переключение на нейрон следующего порядка. Далее по сети нейронов сигнал распространяется в восходящем направлении. Для большинства сенсорных систем, кроме обонятельной, предпоследний нейрон лежит в специфических ядрах таламуса. Отсюда информация поступает в соответствующие проекционные и ассоциативные зоны коры, где в обработку сигнала включаются процессы памяти. Было замечено, что в первичные, проекционные зоны коры импульсы от рецепторов попадают кратчайшим путем, в то время как активация ассоциативных зон происходит несколько позже ввиду вовлечения в этот процесс полисинаптических нервных сетей (см. рис. 3.45).
Рис. 3.45.
Рис. 3.45. Система связей между полями коры мозга человека (по Полякову):
I — первичные (центральные) поля;II – вторичные (периферические) поля;III — третичные (ассоциативные) поля (зоны перекрытия анализаторов).Жирными линиями выделены: система проекционных (корково-подкорковых) связей коры; система проекционно-ассоциативных связей коры; система ассоциативных связей коры.1 — рецептор;2 – эффектор;3 – нейрон чувствительного ганглия;4 – двигательный нейрон;5 –6 – переключательные нейроны спинного мозга и ствола;7–10 – переключательные нейроны подкорковых образований;11, 14 – афферентное волокно из подкорки;13 – пирамида V слоя;16 и 18 – пирамиды слоя III; 12, 15, 17 – звездчатые клетки коры.
Волокна сознательной общей чувствительности
Этот путь проводит через себя все виды общей чувствительности от головы до шеи. Рецепторы начинают свой путь в мышцах и коже, проводят импульсы к чувствительным ганглиям и переходят в ядра тройничного нерва. Далее путь переходит на зрительные бугры, а потом распространяется на клетки постцентральной извилины. При этом включается три основных тракта:
- таламокорковый;
- ганглиоядерный;
- ядерно-таламический.
Волокна сознательной проприоцептивной чувствительности
Этот путь берет свое начало своими рецепторами в сухожилиях, надкостнице, мышцах и связках, а также в суставных сумках. При этом проводится полная информация о вибрациях, положении тела, степени расслабления и сокращения мышц, давлении и весе. Нейроны данного пути располагаются в спинномозговых узлах, ядрах клиновидного и тонкого бугорков продолговатого мозга, зрительного бугра промежуточного мозга, в котором затем начинается переключение импульсов. Информация анализируется и заканчивает свой путь в центральной извилине коры большого полушария мозга. Этот путь включает в себя три тракта:
- Таламокорковый, который заканчивается в проекционном центре, то есть в центральной извилине головного мозга.
- Тонкий и клиновидный пучки, проходящие в заднем канатике спинного мозга.
- Бульбарно-таламический тракт, проходящий в покрышке ствола головного мозга.
ЧУВСТВИТЕЛЬНЫЕ (афферентные) ПРОВОДЯЩИЕ ПУТИ
IV. проводящие пути головного и спинного мозга
Проводящие пути –цепь нейронов, соединяющих функционально однородные участки серого вещества в ЦНС, занимающих в белом и сером веществе головного и спинного мозга определенное место и проводящих одинаковый импульс.
Проводящие пути являются частью сложных рефлекторных дуг, которые соединяют между собой различные отделы центральной нервной системы и обеспечивают двухстороннюю функциональную связь между отдельными структурами головного и спинного мозга. Они отличаются многочисленностью, сложностью строения и надежностью функционирования.
В зависимости от величины, формы и направления нервного импульса проводящие пути получают название: путь (tractus),
пучок
(fasciculus),
волокна
(fibrae),
спайка
(commissura),
петля
(lemniscus)
или лучистость
(radiatio).
Все проводящие пути ЦНС подразделяют на три группы:
1. Проекционные.
2. Комиссуральные.
3. Ассоциативные.
В процессе становления проводящей системы в онтогенезе первоначально формируются проекционные пути, а затем комиссуральные и ассоциативные.
Ассоциативные проводящие пути соединяют участи коры в пределах одного полушария.
Различают:
А. Короткие волокна, дугообразные волокна, fibrae arcuatae cerebri,
которые соединяют корковые поля соседних извилин.
Б. Длинные волокна, которые соединяют корковые поля отдаленных извилин (верхний продольный пучок, fasciculus longitudinalis superior,
нижний продольный пучок,
fasciculus longitudinalis inferior
, пояс,
cingulum
, крючковидный пучок,
fasciculus uncinatus
).
Коммиссуральпые пути соединяют симметричные части правого и левого полушарий. К ним принадлежат:
1. Мозолистое тело, corpus callosum.
2. Передняя спайка, commissura anterior.
Относится к обонятельному мозгу.
3. Спайка свода, commissura hippocampi (fornicis).
Соединяет корковые поля гиппокампа правого и левого полушарий.
4. Задняя спайка, comissura posterior.
5. Спайка поводков, comissura habenulare
.
6. Межталамическое сращение, adhesio intertalamica
.
Проекционные путисоединяют кору с подкорковыми образованиями головного мозга и спинным мозгом.
Проекционные проводящие пути связывают кору головного мозга с его нижележащими отделами (короткие проводящие пути) и со спинным мозгом (длинные проводящие пути).
По направлению проведения нервного импульса проекционные пути подразделяют на две группы:
• афферентные(восходящие, центростремительные, чувствительные), которые проводят нервный импульс от рецепторов, воспринимающих информацию из внешнего мира или внутренней среды организма к различным отделам головного мозга и к коре полушарий;
• эфферентные(нисходящие, центробежные, двигательные), передающие импульс от коры головною мозга и других его отделов на периферию.
ЧУВСТВИТЕЛЬНЫЕ (афферентные) ПРОВОДЯЩИЕ ПУТИ
Под чувствительностью понимают способность организма воспринимать воздействия раздражителей внешней и внутренней среды.
Вид чувствительности определяется характером рецептора, воспринимающего раздражение. Среди рецепторов различают экстерорецепторы(тактильная, болевая, температурная),располагающиеся в коже и слизистых оболочках.; проприорецепторы(мышечно-суставная, вибрационная, чувство давления и веса),которые находятся в мышцах, сухожилиях, связках, суставных капсулах; и висцерорецепторы(чувствительность внутренних органов и сосудов),расположенные в различных внутренних органах и сосудах.
В зависимости от вида чувствительности афферентные проводящие пути подразделяют на:
• экстероцептивные (контактные – общей чувствиельности и дистантные – видовой чувствительности,
• проприоцептивные (корковые – сознательные и мозжечковые – бессознательные),
• интероцептивные.
Те из них, которые заканчиваются в коре полушарий большого мозга, получили название проводников сознательной чувствительности, или проводящих путей коркового направления.
Для чувствительных путей коркового направления характерно:
1. Наличие 3-х нейронов.
2. I-е нейроны представлены псевдоуниполярными нейронами, тела которых расположены в спинно-мозговом узле, а периферические отростки образуют рецепторы.
3. Тела II-х нейронов располагаются в ядрах спинного или продолговатого мозга.
4. Аксоны II-х нейронов образуют (как правило) перекрест.
5. Большинство аксонов II-х нейронов идут к зрительному бугру в составе медиальной петли.
6. Волокна этих путей располагаются, как правило, в дорсальных отделах ствола мозга.
7. Тела III-х нейронов располагаются в латеральных отделах зрительного бугра.
8. Все они проходят в средней трети заднего бедра внутренней капсулы.
9. Аксоны III-х нейронов по пути в кору образуют или проходят в составе лучистого венца.
Спиномозговые волокна
Афферентные пути спинного мозга образуются при помощи аксонов, или, как их еще называют по-другому, окончания нейронов. Аксоны находятся только в спинном мозге и не выходят за его пределы, а также создают связь между всеми сегментами органа. Атомическое строение данных волокон состоит в том, что длина аксонов довольно большая и соединяется с другими нервными окончаниями. От рецепторов к центральной нервной системе за счет афферентных проводящих путей спинного и головного мозга проводятся нервные сигналы. Все нервные волокна, расположенные по всей длине спинного мозга, участвуют в данном процессе. Сигнал к органам проводится от разных отделов ЦНС и между нейронами. Беспрепятственное прохождение сигнала от периферии к центральной нервной системе достигается при помощи проводящих путей спинного мозга.
Задний и передний спиномозжечковые тракты
Афферентные пути мозжечка относятся к бессознательным и берут свое начало в боковом канатике спинного мозга, а оттуда уже проводят информацию о том, в каком состоянии находятся органы опорно-двигательного аппарата. Передний спиномозжечковый тракт поступает в мозжечок через верхнюю ножку, в связи с этим он проходит через покрышку продолговатого мозга, среднего мозга и моста. Задний спиномозжечковый тракт проходит в продолговатом мозге и проникает через нижнюю ножку.
Эти два тракта передают в мозжечок информацию от связок, суставных сумок, рецепторов мышц, сухожилий, надкостницу. Именно они отвечают за поддержание равновесия и координацию движений человека, поэтому роль их в организме очень важна.
Слуховые волокна
Данный путь проводит информацию от рецепторов Кортиева органа, который располагается во внутреннем ухе. Нервные импульсы поступают в мост, содержащий в себе слуховые ядра по волокнам преддверно-улиткового нерва. Через слуховые ядра информация передается на ядра трапециевидного тела. После этого импульсы поступают на подкорковые центры слуха, которые включают в себя зрительный бугор, нижние холмики и коленчатые медиальные тела.
В среднем мозге на эти слуховые раздражители возникают возвратные реакции, при этом афферентные слуховые пути переключаются в ядра таламуса, в которых происходит оценка слуховых раздражителей – они отвечают за движения, происходящие непроизвольно: ходьба, бег. От коленчатых тел начинает исходить слуховая лучистость – этот тракт проводит импульсы от внутренней капсулы до проекционного центра слуха. Только здесь уже начинает происходить оценка звуков. В задней части височной извилины располагается ассоциативный слуховой центр. Именно в нем все звуки начинают восприниматься как слова.
Некоторые закономерности строения афферентных
Проекционных проводящих путей.
1. Начало каждого пути представлено рецепторами, заложенными в коже, подкожной ткани или глубоких частях тела.
2. Первый нейрон у всех афферентных путей находится вне центральной нервной системы, в спинномозговых ганглиях.
3. Второй нейрон локализуется в ядрах спинного или продолговатого мозга.
4. Все восходящие пути проходят в покрышке мозгового ствола.
5. Третий нейрон у путей, направляющихся в кору полушарий большого мозга, располагается в ядрах таламуса, а у мозжечковых путей — в коре мозжечка.
6. Пути, приносящие импульсы в кору большого мозга, имеют один перекрест, совершаемый отростками 2-го нейрона; благодаря этому каждая половина тела проецируется на противоположное полушарие большого мозга.
7. Мозжечковые пути или не имеют ни одного перекреста, или перекрещиваются дважды, так что каждая половина тела проецируется на кору одноименной половины мозжечка.
8. Пути, соединяющие мозжечок с корой полушарий большого мозга, являются перекрещенными.
Эфферентные проводящие пути.
Нисходящие проекционные проводящие пути (эффекторные, эфферентные) проводят импульсы от коры, подкорковых центров к нижележащим отделам, к ядрам мозгового ствола и двигательным ядрам спинного мозга. Эти пути подразделяются на 2 группы: 1) пирамидный путь и 2) экстрапирамидные пути.
Пирамидный путь
Пирамидный путь (tractus pyramidalis)
связывает нейроны двигательной области коры непосредственно с двигательными ядрами спинного мозга и черепных нервов. Началом пути являются большие пирамидные нейроны (клетки Беца) (1-й нейрон), расположенные во внутреннем пирамидном слое коры предцентральной извилины (первичное корковое поле 4).
Пирамидный путь подразделяется на корково-спинномозговой (fibrae corticospinales)
и
корково-ядерный (fibrae corticonucleares)
. Последний будет рассмотрен при описании проводящих путей черепных нервов.
Корково-спинномозговой путь составлен аксонами больших пирамидных нейронов, расположенных в верхней и средней третях предцентральной извилины. Они проходят через передний отдел задней ножки внутренней капсулы, среднюю часть основания ножки большого мозга, основание моста и пирамиду продолговатого мозга. На границе со спинным мозгом происходит неполный перекрест корково-спинномозгового пути (decussatio pyramidum)
. Большая часть волокон, перейдя на противоположную сторону, образует
латеральный корково-спинномозговой путь (tractus corticospinalis lateralis)
, остальные идут в
Рис. 7. Пирамидный путь.
составе переднего корково-спинномозгового пути (tractus corticospinalis anterior)
своей стороны и перекрещиваются в спинном мозге посегментно, проходя через белую спайку. Они преимущественно оканчиваются в промежуточном сером веществе, образуя синапсы со вставочными нейронами (2-й нейрон), которые передают импульсы мотонейронам ядер переднего рога (3-й ней-рон). Аксоны двигательных клеток передних рогов выходят из спинного мозга в передних корешках и затем идут в составе спинномозговых нервов к скелетным мышцам, обеспечивая их двигательную иннервацию.
Через вставочный нейроциты осуществляется связь клеток коры полушарий мозга с мотонейронами, иннервирующими мускулатуру шеи, туловища и проксимальных отделов конечностей. Волокна латерального пирамидного пути заканчиваются в своем большинстве на вставочных нейроцитах спинного мозга.
Мотонейроны спинного мозга, иннервирующие мышцы предплечья и кисти, имеют непосредственные связи с клетками коры. На них (а не на вставочных нейронах) заканчиваются корково-спинномозговые волокна, проходящие главным образом в составе переднего пирамидного тракта. Этим достигается прямая, более совершенная корковая регуляция мышц, которые характеризуются очень точными, строго дифференцированными движениями.
Пирамидный путь главным образом передает мышцам сигналы произвольных движений, регулируемых корой большого мозга. При его перерыве наступает паралич мышц своей или противоположной стороны в зависимости от уровня повреждения. Особенно сильно страдают при поражениях пирамидных путей тонко дифференцированные движения верхних конечностей, мускулатура которых имеет перекрестную иннервацию. Мышцы нижних конечностей и особенно мышцы туловища иннервируются наряду с перекрещенными, также и неперекрещенными корково-спинномозговыми волокнами, поэтому их функция нарушается в меньшей степени.
Однако пирамидный путь связывает кору не только с двигательными нейроцитами спинного мозга, но и с другими структурами головного и спинного мозга. Его волокна или их коллатерали оканчиваются на клетках красного ядра, собственных ядрах моста, на клетках ретикулярной формации ствола мозга.
Экстрапирамидные пути.
Экстрапирамидная система представлена многозвенными нисходящими путями, посредством которых осуществляется регуляция непроизвольных движений, автоматических двигательных актов, мышечного тонуса, а также движений, выражающих эмоции (улыбка, смех, плач и т. п.).
Нейроны внутреннего пирамидного слоя коры лобной доли (поле 6) (1-нейрон) посылают корково-стриальные волокна в новую часть полосатого тела, представленную хвостатым ядром и скорлупой. Здесь локализуется 2-й нейрон экстрапирамидных путей, отростки которого идут в древнюю часть полосатого тела — бледный шар (стриато-паллидарные волокна). Нервные клетки бледного шара представляют собой 3-й нейрон, их аксоны идут в составе чечевицеобразной петли (ansa lenticularis)
к различным ядрам мозгового ствола — субталамическому ядру, черному веществу, ядрам верхних холмиков, красному ядру, латеральному преддверному ядру,
Рис. 9. Корково-мосто-мозжечково-спинномозговые пути.
оливному ядру, ретикулярным ядрам. В названных ядрах заложен 4-й нейрон, который дает начало нисходящим путям, передающим сигналы двигательным ядрам черепных нервов и спинного мозга покрышечно-спинномозговому (tractus tectospinalis),
красноядерно-спинномозговому (tractus rubrospinalis),преддверно-спинномозговому (tractus vestibulospinalis),оливо-спинномозговому (tractus olivospinalis),ретикулярно-спинномозговому (fasciculi reticulospinales)
. Наиболее хорошо развиты у человека красноядерно-спинномозговой и ретикулярно-спинномозговой пути, остальные представлены немногочисленными волокнами и прослеживаются обычно лишь в шейных сегментах спинного мозга. Двигательные клетки ядер черепных нервов и передних рогов спинного мозга образуют 5-й нейрон экстрапирамидных путей, посылающий импульсы скелетным мышцам.
К экстрапирамидным путям относят также систему волокон, связывающих кору большого мозга с мозжечком. Эта система состоит из двух звеньев. Первым являются корково-мостовые пути (tractus corticopontini)
. Начало им дают клетки внутреннего пирамидного слоя вторичных корковых полей лобной (поля 6 и 8), теменной, затылочной, височной долей; соответственно этому различают лобно-, теменно-, затылочно- и височно-мостовой пути. Все они проходят через переднюю ножку внутренней капсулы, основание ножки большого мозга (ее медиальный и латеральный отделы) и оканчиваются в собственных ядрах моста, где заложен 2-й нейрон
(nuclei pontis)
. Аксоны клеток, образующих ядра моста, по средним мозжечковым ножкам в составе
tractus pontocerebellaris
переходят на противоположную сторону (1-й перекрест) и заканчиваются в коре полушарий мозжечка (новый мозжечок). Грушевидные клетки коры мозжечка принимаются за 3-й нейрон. Посылаемые ими импульсы поступают в зубчатое ядро (4-й нейрон). Отсюда происходит передача импульсов по
зубчато-красноядерному пути (tractus dentatorubralis)
через верхние мозжечковые ножки к красному ядру (5-й нейрон). В покрышке среднего мозга происходит 2-й перекрест
(decussatio pedunculorum cerebellarium superiorum)
. От красного ядра начинается
красноядерно-спинномозговой путь (tractus rubrospinalis)
, который после перекреста
(decussatio ventralis tegmenti)
идет к ядрам передних рогов спинного мозга и двигательным ядрам черепных нервов (6-й нейрон). Отсюда в составе спинномозговых и черепных нервов импульсы поступают в мускулатуру.
Рис. 8. Экстрапирамидные пути.
Посредством корково-мостового и мосто-мозжечкового путей и восходящих эфферентных путей мозжечка осуществляется кольцевое взаимодействие между корой большого мозга и мозжечка, необходимое для регуляции и координирования различных двигательных актов. Мозжечок получает из коры большого мозга как бы копии команд, посылаемых по пирамидным и экстрапирамидным путям, сопоставляет их с сигнализацией поступающей от проприоцепторов и вестибулярного аппарата, и направляет переработанную информацию высшим двигательным центрам коры.
Вкусовые анализаторы
Импульсы афферентного пути вкусовых анализаторов развиваются от рецепторов корня языка, входящих в состав языкоглоточных нервов и располагающихся на языке, которые входят в состав лицевого нерва. Импульсы от них поступают в продолговатый мозг, а потом к ядрам лицевого и языкоглоточного нерва. Наименьшая часть всей получаемой информации от этих импульсов доставляется в мозжечок, образуя тем самым ядерно-мозжечковый путь, и обеспечивает рефлекторную регуляцию тонуса мышц языка, головы и глотки. Большая часть информации поступает в зрительные бугры, после чего импульсы достигают крючка височной доли, в котором происходит их сознательный анализ.
Леся Украинка
— псевдоним Ларисы Петровны Косач, знаменитой украинской писательницы и поэтессы.
Леся Украинка последовательный и энергичный борец за образование украинского народа, за его консолидацию в политическую нацию. Вся совокупность ее произведений — это образ идеальной земли, земного рая, называемого.
Леся Украинка родилась 25 февраля 1871 года в г. Новоград–Волынском
, в той части Украины, которая входила в состав Российской империи. В семье не чуждой высоких духовных интересов:
мать
— писательница, творившая под псевдонимом: Олена Пчилка (её поэзию и рассказы на родном языке для детей, хорошо знали на Украине),
отец
— высокообразованный помещик, очень любивший литературу и живопись. В доме Косачей часто собирались писатели, художники и музыканты, устраивались вечера и домашние концерты. Дядя Леси (так ее звали в семье, и это домашнее имя стало ее литературным псевдонимом.) — Михаил Драгоманов, впоследствии дружески опекавший племянницу и всяческий помогавший ей, — был известным ученым, общественным деятелем, подолгу жил за границей во Франции и Болгарии. Он водил знакомство с Иваном Сергеевичем Тургеневым, Виктором Гюго, был в курсе всех новейших литературных и политических событий и часто пополнял библиотеку племянницы посылками из — за границы.
Писать и печататься Леся Украинка начала в детском возрасте (12 лет).
К ее ранним произведениям относится украинский перевод «Вечеров на хуторе» Гоголя (в соавторстве с братом). Печаталась Украинка в заграничных журналах «Зоря», «Жите і слово», «Літературно-науковий вістник».
Необычайно талантливая, восприимчивая, ранимая, с глубоким, истинным, музыкальным дарованием, (она начала играть и сочинять маленькие музыкальные пьесы с пяти лет
!) в возрасте восьми лет написавшая первое стихотворение, Леся в 1881 году неожиданно тяжело заболела. Ее мучила нестерпимая боль в правой ноге. Вначале решили, что у нее острый ревматизм, лечили ваннами, мазями, травами, но все было бесполезно. Боль перешла в руки. Врачи, наконец, смогли определить, что это —
туберкулез кости
. На музыкальной карьере Леси был поставлен крест. После первой, сложной, но крайне неудачной, операции рука осталось искалеченной! Тогда -то в глазах хрупкой девочки впервые появилась печаль. Она и в дальнейшем, словно легкое покрывало, будет окутывать все ее творчество. Отныне много месяцев в году девочка должна лежать в постели, не делать резких движений, все время испытывать мучительную боль…
Леся Украинка всегда и во всем пыталась отыскать радость, даже в малом! В ней жил неукротимый дух. Самозабвенно, ночами, изучала языки: болгарский, испанский, латинский, древнегреческий, итальянский, польский, немецкий, не говоря об английском и французском, географию, историю Востока и восточных культур, историю искусства и религий, а для своих младших сестер в 19 летнем возрасте написала учебник: «Древняя история восточных народов».
В 1893 году, во Львове, (Западная Украина) вышла тоненькая книжечка ее стихов названная: «На крыльях песни» и тепло встреченная критикой и публикой. Книга быстро сделала ее популярной.
В 1898 году в Запорожье Лариса Петровна познакомилась с Сергеем Константиновичем Мержинским, общественным деятелем, выпускником Киевского университета святого Владимира. Мержинский жил некоторое время в Ялте, проходя лечение от туберкулёза. Четыре года спустя (в 1901 году) Леся едет в зимний Минск к смертельно больному возлюбленному. В тяжелые зимние месяцы рождается одна из сильнейших её драм — «Одержима», Сергей Мержинский умирает, и Лариса Петровна навсегда надевает чёрные траурные одежды.
Тридцати шести лет от роду, она вновь полюбила. Человека, который на ее чувства ответил не менее искренней и глубокой привязанностью — Климента Квитка, ученого музыковеда- фольклориста, собирателя народных преданий и песен Мать Леси снова была яростно против всяческих отношений дочери «с каким -то нищим», как она презрительно называла Климента — человека мягкого по характеру, замкнутого, стеснительного, пережившего в детстве глубокую личную драму -он рос в приемной семье. Но Квитка так страстно привязался к тоненькой, больной женщине с большими печальными глазами, понимающей его с полуслова, что наотрез отказался ее покинуть! И, несмотря на весь гнев и мрачные прогнозы будущего молодых, Ольга Петровна была вынуждена согласиться на брак дочери. Молодые приняли решение: отказались от помощи родителей. Все деньги, необходимые на лечение тяжелобольной жены Климент зарабатывал сам. Продавали все, что можно было продать: вещи, нехитрый скарб, кухонную утварь. Дорожили только библиотекой.
Леся лечилась в Египте и Греции, в Германии и Австрии. Все было бесполезно. К обострившемуся процессу костного туберкулеза прибавилась неизлечимая болезнь почек. Она скончалась в г. Сурами (Грузия) первого августа 1913 года
. Улетела «на крыльях песни». Осуществилась ее давняя мечта: она всегда хотела потрогать руками облака…
Литературная деятельность Леси Украинки протекала в эпоху империализма и первой русской революции. В украинской литературе того времени определились натуралистическое, декадентско-символистское и революционно-демократическое течения. К последнему, резко противопоставлявшемуся всей буржуазно-националистической литературе, примыкали Грабовский П., Коцюбинский М. и Тесленко. Л. Украинка относилась отрицательно как к декадентам, так и к натуралистам. Отходя от либералов, она все более и более проникалась революционными идеями, приближаясь к революционным демократам.
Творческий метод Леси Украинки более близок к романтизму, чем к реализму, но в основе он резко отличался от метода декадентствовавших украинских неоромантиков, в особенности символистов, прежде всего своей идейной и тематической направленностью, а также художественными средствами. В отличие от многих романтиков-декадентов Леся Украинка не идеализировала прошлого, хотя и создавала свои образы на историческом материале; страстно ненавидя гнетущую действительность царской России, она не предавалась пессимизму, не впадала в отчаяние, а наоборот, звала к борьбе за полное освобождение от всякого гнета и уничтожение эксплуатации человека человеком. Романтизм Леси Украинки был пронизан революционной настроенностью.
Произведения, написанные Лесей Украинкой:
- сборник поэзий «На крыльях песен» («На крилах пісень», 1893 год);
- сборник поэзий «Думы и мечты» («Думи і мрії», 1899 год);
- сборник поэзий «Отклики» («Відгуки», 1902 год);
- Драма-феерия «Лесная песня» («Лісова пісня», 1911 год).
Источники информации:
- Википедия: Леся Украинка;
- Леся Украинка: биографическая справка;
- Леся Украина — биография;
- Леся Украинка — краткая биография.
Зрительные анализаторы
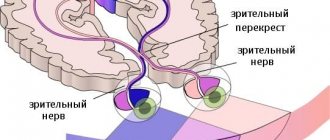
Афферентные проводящие пути ЦНС зрительного анализатора начинаются от колбочек и палочек сетчатки глазного яблока. Импульсы поступают в зрительный перекресток в составе зрительных нервов, а затем по тракту направляются к подкорковым центрам головного мозга, то есть в зрительный бугор, коленчатые латеральные тела и верхние холмики, расположенные в средней части мозга.
В среднем мозге на эти раздражители возникает ответная реакция, при этом в ядрах таламуса начинается бессознательная оценка импульсов, которые обеспечивают непроизвольные движения, воспроизводимыми человеком. Основными такими бессознательными движениями являются бег и ходьба. В проекционном центре зрения или в шпорную борозду затылочной доли головного мозга импульсы поступают по зрительной лучистости от коленчатых тел, находящихся в составе внутренней капсулы, после чего начинается полный анализ поступающих данных. В коре, которая прилегает к шпорной борозде, находит свое место расположения центральная часть, отвечающая за зрительную память, которую еще называют ассоциативным зрительным центром.
Обонятельный анализатор
Афферентный путь обонятельного анализатора берет свое начало от рецепторов слизистой оболочки, локализующейся в верхней части носового хода. После этого импульсы направляются к аксонам обонятельных луковиц, и протекают они по волокнам обонятельных нервов. Затем импульсы направляются к проекционному центру обоняния, который находится в районе парагиппокампальной извилины и крючка. Следуют эти импульсы по тракту до коры височной доли головного мозга. К подкорковым центрам, которые располагаются в средней и промежуточной части мозга, направляется большая часть информации, получаемая от обонятельных рецепторов. Подкорковые центры мозга в ответ на обонятельные раздражители обеспечивают рефлекторную регулировку тонуса мышц.
Исходя из этого, можно определить, что основная особенность обонятельных рецепторов заключается в том, что нервные импульсы изначально поступают в кору полушарий головного мозга, а не в подкорковые центры обоняния. В связи с этим, человек сначала чувствует запах, затем начинает оценивать его и только после этого в мозге формируется бессознательная окраска раздражителя на эмоциональном уровне. На весь процесс при этом затрачиваются всего лишь доли секунды.
ЛЕКЦИИ ПО НЕЙРОПСИХОЛОГИИ: Лекция № 4 5/5 (2)
Лекция 4. Нарушения произвольных движений и действий.
- Двигательный анализатор: афферентные и эфферентные механизмы.
- Элементарные двигательные расстройства.
- Проблема апраксий.
Произвольные движения и действия относятся к числу наиболее сложных психических функций человека. Прозвольные действия — совокупность произвольных движений, объединенных единой целью. Произвольные движения и действия условно-рефлекторные по своему происхождению, формируются полностью прижизненно.
Современные психологические представления о произвольных движениях сложились в борьбе как с идеалистическим подходом, так и с вульгарно-материалистическими, механистическими концепциями (бихевиоризмом и др.). Важный вклад в современное понимание произвольных движений внесли отечественные физиологи (И. М. Сеченов, И. П. Павлов, Н. А. Бернштейн, П. К. Анохин и др.) и психологи (Л. С. Выготский, А. Н. Леонтьев, А. Р. Лурия,). Произвольные движения и действия могут быть как самостоятельными двигательными актами, так и средствами, с помощью которых реализуются самые различные формы поведения.
Произвольные движения входят в состав устной и письменной речи, а также многих других высших психических функций. С физиологической точки зрения к произвольным движениям относятся движения поперечно-полосатой мускулатуры рук, лица ног, всего туловища, т. е. обширнейший класс движений.
Современная физиология располагает разнообразными сведениями относительно большой сложности как афферентного, так и эфферентного механизмов произвольных движений. Афферентный механизм произвольных движений afferentis — «приносящий» Произвольные движения — это сложно афферентированные системы, включающие разные виды афферентации, среди которых базальной является кинестетическая афферентация.
Любое движение — сложная многоуровневая система, где каждый уровень (или определенные анатомические структуры) характеризуется «ведущей афферентацией» и собственным набором регулируемых движений.
Выделяют пять уровней регуляции движений: 1. рубро-спинальный; 2. таламо-паллидарный; 3. пирамидно-стриальный; 4. теменно-премоторный; 5. корковый «символический».
Все эти уровни объединяют непроизвольные и произвольные движения в единую систему. Первый и второй уровни ответственны за регуляцию непроизвольных движений (к ним относятся движения гладкой мускулатуры, тремор, тонус, и др.).
Третий-пятый уровни связаны с регуляцией произвольных двигательных актов, в которых участвуют как движения всего туловища, так и отдельных частей тела: рук, лица, речевого аппарата и т. д.
Таким образом, произвольные движения — это целый набор различных двигательных актов, регулируемых разными уровнями (структурами) нервной системы и управляемых разного рода афферентными импульсами (и различной «ведущей афферентацией»).
Поражение любого из перечисленных уровней ведет к нарушениям движений данного уровня, а также тех двигательных актов, куда эти движения включаются как «фоновые». Тип афферентации, а также соответствующие анатомические структуры являются критериями для выделения класса движений (это относится как к произвольным, так и к непроизвольным движениям). Иными словами, афферентация является важнейшим фактором, определяющим тип движения.
А. Р. Лурия, анализируя наблюдения над больными с локальными поражениями головного мозга, описал конкретный состав корковых зон, участвующих в мозговой организации произвольных двигательных актов, включив в понятие «двигательный организатор» не только моторные, но и сенсорные, и ассоциативные корковые поля.
Лурия отмечал, что помимо собственно двигательных, моторных зон коры больших полушарий в корковое звено двигательного анализатора следует включать и многие другие зоны коры, а именно:
- постцентральную теменную кору, обеспечивающую анализ кожно-кинестетической афферентации, поступающей от органов движения; 2. задние затылочные и теменно-затылочные отделы коры больших полушарий, которые обеспечивают регуляцию движений с помощью зрительной афферентации, а также ответственны за пространственную организацию движений; 3. височную кору (прежде всего левого полушария), обеспечивающую не только слухоречевую афферентацию речевой моторики, но и участвующую во всех «оречевленных» (внешней и внутренней речью) двигательных актах; 4. передние отделы коры больших полушарий (премоторную и префронтальную кору), с помощью которых осуществляются программирование движений, организация движений во времени и контроль за выполнением программы.
Следовательно, произвольные движения человека обеспечиваются самыми различными видами афферентации, и поэтому в корковое звено двигательного анализатора, следует включать почти всю кору больших полушарий. В целом, в современной нейрофизиологии и нейропсихологии сложились представления о том, что произвольные движения — это очень сложно афферентированные системы, которые реализуются при участии почти всей коры больших полушарий.
Эфферентные механизмы произвольных движений (эфф – выносящий) К эфферентным (исполнительным) механизмам произвольных движений и действий относятся, как известно, две взаимосвязанные, но в определенной степени автономные эфферентные системы: пирамидная и экстрапирамидная, корковые отделы которых составляют единую сенсомоторную зону коры.
Пирамидная система — это основной механизм, реализующий произвольные движения. Начинается от моторных клеток Беца, находящихся в V слое моторной коры (4-е поле), продолжается в виде корково-спинномозгового, или пирамидного, тракта, который переходит на противоположную сторону в области пирамид и заканчивается на мотонейронах спинного мозга (на 2-м нейроне пирамидного пути), иннервирующих соответствующую группу мышц. К этим представлениям о пирамидной системе как об основном эфферентном механизме произвольных движений добавились новые данные:
Во-первых, не только 4-е поле является моторным. Моторные клетки пирамидного типа обнаружены в 6-м и 8-м полях прецентральной коры, и во 2, 1 и 3-м полях постцентральной коры. Следовательно, пирамидный путь начинается не только от 4-го поля, как это предполагалось ранее, а со значительно больших площадей коры больших полушарий. Только 40 % всех волокон пирамидного пути начинается в 4-м поле, около 20 % — в постцентральной извилине; остальные — в премоторной зоне коры больших полушарий.
Во-вторых, установлено, что пирамидный путь содержит волокна различного типа (по диаметру и степени миелинизации). Хорошо миелинизированные волокна составляют не более 10 % всех пирамидных волокон, которые идут от коры к периферии.
В-третьих, если ранее предполагалось, что существует единый пирамидный, или кортико-спинальный, путь (латеральный), который идет с перекрестом в зоне пирамид от коры больших полушарий к мотонейронам спинного мозга, то в настоящее время выделен другой кортико-спинальный путь (вентральный), идущий без перекреста в составе пирамид на той же стороне. В-четвертых, пирамидный путь оканчивается не непосредственно на мотонейронах, расположенных в передних рогах спинного мозга, как считалось ранее, а главным образом на промежуточных (или вставочных) нейронах, с помощью которых модулируется возбудимость основных мотонейронов и тем самым оказывается воздействие на конечный результат — произвольные движения. Все эти данные свидетельствуют о сложности пирамидной системы как исполнительного механизма.
Помимо моторных зон коры больших полушарий, раздражение которых вызывает движения, существуют и такие зоны коры, раздражение которых прекращает уже начавшееся движение (подавляющие области коры).
В коре больших полушарий имеются еще особые адверзивные зоны. Это области коры, хорошо известные нейрохирургам и невропатологам. Их раздражение (электрическим током или болезненным процессом) вызывает адверзивные эпилептические припадки (начинающиеся с адверзии — поворота туловища, глаз, головы, рук и ног в сторону, противоположную расположению возбуждающего агента). Эпилепсия, протекающая с припадками этого типа, известна как «эпилепсия Джексона».
Существуют две адверзивные зоны коры: премоторная и теменно-затылочная. (поля 6, 8 и 19-е). Эти поля коры участвуют в реакциях, связанных с вниманием к раздражителю, т. е. в организации сложных двигательных актов, опосредующих внимание к определенному стимулу.
Экстрапирамидная система — второй эфферентный механизм реализации произвольных движений и действий. Экстрапирамидной системой называются все двигательные пути, которые не проходят через пирамиды продолговатого мозга.
Экстрапирамидная система более древняя, чем пирамидная. Она очень сложна по своему составу. В экстрапирамидной системе различают корковый и подкорковый отделы. К корковому отделу экстрапирамидной системы относятся те же поля, которые входят в корковое ядро двигательного анализатора. Это 6-е и 8-е, а также 1-е и 2-е поля, т. е. сенсомоторная область коры.
Строение подкоркового отдела экстрапирамидной системы довольно сложное. Он состоит из целого ряда образований. Конечной инстанцией экстрапирамидных влияний являются те же мотонейроны спинного мозга, к которым адресуются импульсы и пирамидной системы.
!!!! Четкая анатомическая граница между пирамидной и экстрапирамидной системами отсутствует. Они обособлены анатомически только на участке пирамид, в продолговатом мозге. Однако функциональные различия между этими системами достаточно отчетливы. И особенно ясно они проявляются в клинике локальных поражений головного мозга.
Нарушения двигательных функций, возникающие при различных локальных поражениях мозга, можно подразделить на относительно элементарные, связанные с поражением исполнительных, эфферентных механизмов движений, и более сложные, распространяющиеся на произвольные движения и действия и связанные преимущественно с поражением афферентных механизмов двигательных актов.
Относительно элементарные двигательные расстройства возникают при поражении подкорковых звеньев пирамидной и экстрапирамидной систем. При поражении коркового звена пирамидной системы (4-го поля), расположенного в прецентральной области, наблюдаются двигательные расстройства в виде парезов или параличей определенной группы мышц: руки, ноги или туловища на стороне, противоположной поражению. Для поражения 4-го поля характерен вялый паралич (когда мышцы не сопротивляются пассивному движению), протекающий на фоне снижения мышечного тонуса. Но при очагах, расположенных кпереди от 4-го поля (в 6-м и 8-м полях коры), возникает картина спастического паралича, т. е. выпадений соответствующих движений на фоне повышения мышечного тонуса. Явления парезов вместе с чувствительными расстройствами характерны и для поражения постцентральных отделов коры. Эти нарушения двигательных функций подробно изучаются неврологией. Наряду с этими неврологическими симптомами поражение коркового звена экстрапирамидной системы дает также нарушения сложных произвольных движений, о которых речь будет идти ниже.
При поражении пирамидных путей в подкорковых областях мозга (например, в зоне внутренней капсулы) возникает полное выпадение движений (паралич) на противоположной стороне. Полное одностороннее выпадение движений руки и ноги (гемиплегия) появляется при грубых очагах. Чаще в клинике локальных поражений мозга наблюдаются явления частичного снижения двигательных функций на одной стороне (гемипарезы).
При пересечении пирамидного пути в зоне пирамид — единственной зоне, где пирамидный и экстрапирамидный пути анатомически обособлены, — произвольные движения реализуются только с помощью экстрапирамидной системы. При пересечении пирамид движения становятся менее дискретными (т. е. более грубыми). В захватывании предметов начинает участвовать большее количество мышц. Кроме того, в движенииях, где требуется участие всех мышц (типа прыжка), что характерно для моторики обезьян (на них проводились операции), исчезают точность и координированность. Подобные нарушения движений протекают на фоне повышения мышечного тонуса. Эти данные свидетельствуют о том, что пирамидная система участвует в организации преимущественно точных, дискретных, пространственно-ориентированных движений и в подавлении мышечного тонуса. Поражение корковых и подкорковых звеньев экстрапирамидной системы приводит к появлению различных двигательных расстройств. Эти расстройства можно подразделить на динамические (т. е. нарушения собственно движений) и статические (т. е. нарушения позы).
При поражении коркового уровня экстрапирамидной системы (6-е и 8-е поля премоторной коры), в контралатеральных конечностях возникают спастические двигательные нарушения. Раздражение 6-го или 8-го полей вызывает повороты головы, глаз и туловища в противоположную сторону (адверзии), а также сложные движения контралатеральных руки или ноги.
Поражение подкорковой стриопаллидарной системы, вызванное различными заболеваниями (паркинсонизмом, болезнью Альцгеймера, Пика, опухолями, кровоизлияниями в область базальных ядер и др.), характеризуется общей неподвижностью, адинамией, трудностями передвижения. Одновременно появляются насильственные движения контралатеральных руки, ноги, головы — гиперкинезы. У таких больных наблюдаются и нарушение тонуса (в виде спастичности, ригидности или гипотонии), составляющего основу позы, и нарушение двигательных актов (в виде усиления тремора — гиперкинезов). Больные теряют возможность себя обслуживать и становятся инвалидами. В настоящее время разработаны хирургические (стереотаксические) и консервативные (фармакологические) методы лечения паркинсонизма. Суть первых состоит в разрушении одной из «больных» экстрапирамидных структур (например, вентролатерального ядра таламуса) и прекращении патологического возбуждения, циркулирующего по замкнутым экстрапирамидным путям. В зависимости от формы паркинсонизма разрушают также либо зону паллидума, либо зону стриатума (в области хвостатого ядра или скорлупы). Зона паллидума (бледный шар) имеет отношение к регуляции медленных плавных движений и к поддержанию тонуса (позы), в то время как зона стриатума (скорлупа и хвостатое ядро) в большей степени связана с физиологическим тремором, переходящим при патологии в гиперкинезы. Избирательное поражение зоны паллидума (более древней части, чем стриатум) может привести к атетозу или хореоатетозу (патологическим волнообразным движениям рук и ног, подергиванию конечностей и т. д.).
Поражение стриопаллидарных образований сопровождается еще одним видом двигательных симптомов — нарушением мимики и пантомимики, т. е. непроизвольных моторных компонентов эмоций. Эти нарушения могут выступать либо в форме амимии (маскообразное лицо) и общей обездвиженности, либо в форме насильственного смеха, плача или насильственной ходьбы, бега (пропульсии). Нередко у этих больных страдает и субъективное ‘переживание эмоций. Наконец, у таких больных нарушаются и физиологические синергии — нормальные сочетанные движения разных двигательных органов (например, размахивание руками при ходьбе), что приводит к неестественности их двигательных актов.
Последствия поражения других структур экстрапирамидной системы изучены в меньшей степени, за исключением, конечно, мозжечка. Мозжечок представляет собой важнейший центр координации различных двигательных актов, «орган равновесия», обеспечивающий целый ряд безусловных моторных актов, связанных со зрительной, слуховой, кожно-кинестетической, вестибулярной афферентацией. Поражение мозжечка сопровождается разнообразными двигательными расстройствами (прежде всего расстройствами координации двигательных актов). Их описание составляет один из хорошо разработанных разделов современной неврологии.
Поражение пирамидных и экстрапирамидных структур спинного мозга сводится к нарушению функций мотонейронов, вследствие чего выпадают (или нарушаются) управляемые ими движения. В зависимости от уровня поражения спинного мозга нарушаются двигательные функции верхних или нижних конечностей (на одной или на обеих сторонах), причем все местные двигательные рефлексы осуществляются, как правило, нормально или даже усиливаются вследствие устранения коркового контроля. Все эти нарушения движений также подробно рассматриваются в курсе неврологии.
Клинические наблюдения за больными, у которых имеется поражение того или иного уровня пирамидной или экстрапирамидной системы, позволили уточнить функции этих систем. Пирамидная система ответственна за регуляцию дискретных, точных движений, полностью подчиненных произвольному контролю и хорошо афферентированных «внешней» афферентацией (зрительной, слуховой). Она управляет комплексными пространственно-организованными движениями, в которых участвует все тело. Пирамидная система регулирует преимущественно фазический тип движений, т. е. движения, точно дозированные во времени и в пространстве.
Экстрапирамидная система управляет в основном непроизвольными компонентами произвольных движений; к ним кроме регуляции тонуса (того фона двигательной активности, на котором разыгрываются фазические кратковременные двигательные акты) относятся: • поддержание позы; • регуляция физиологического тремора; • физиологические синергии; • координация движений; • общая согласованность двигательных актов; • их интеграция; • пластичность тела; • пантомимика; • мимика и т. д.
Экстрапирамидная система управляет также разнообразными двигательными навыками, автоматизмами. В целом экстрапирамидная система менее кортиколизована, чем пирамидная, и регулируемые ею двигательные акты в меньшей степени произвольны, чем движения, регулируемые пирамидной системой. Следует, однако, помнить, что пирамидная и экстрапирамидная системы представляют собой единый эфферентный механизм, разные уровни которого отражают разные этапы эволюции. Пирамидная система, как эволюционно более молодая, является в известной степени «надстройкой» над более древними экстрапирамидными структурами, и ее возникновение у человека обусловлено прежде всего развитием произвольных движений и действий. Нарушения произвольных движений и действий
Нарушения произвольных движений и действий относятся к сложным двигательным расстройствам, которые в первую очередь связаны с поражением коркового уровня двигательных функциональных систем. Этот тип нарушений двигательных функций получил в неврологии и нейропсихологии название апраксий. Под апраксиями понимаются такие нарушения произвольных движений и действий, которые не сопровождаются четкими элементарными двигательными расстройствами — параличами и парезами, явными нарушениями мышечного тонуса и тремора, хотя возможны сочетания сложных и элементарных двигательных расстройств.
Апраксиями обозначают прежде всего нарушения произвольных движений и действий, совершаемых с предметами. Сложности понимания природы апраксий нашли отражение в их классификациях. Наиболее известная классификация, предложенная в свое время Г. Липманном выделяет три формы апраксий: • идеаторную, предполагающую распад «идеи» о движении, его замысла; • кинетическую, связанную с нарушением кинетических «образов» движения; • идеомоторную, в основе которой лежат трудности передачи «идей» о движении в «центры исполнения движений». Первый тип апраксий Г. Липманн связывал с диффузным поражением мозга, второй — с поражением коры в нижней премоторной области, третий — с поражением коры в нижней теменной области.
А. Р. Лурия разработал классификацию апраксий, исходя из общего понимания психологической структуры и мозговой организации произвольного двигательного акта. Он выделил четыре формы апраксий
Первая — кинестетическая апраксия. Эта форма апраксий, возникает при поражении нижних отделов постцентральной области коры больших полушарий. В этих случаях нет четких двигательных дефектов, сила мышц достаточная, однако страдает кинестетическая основа движений. Они становятся недифференцированными, плохо управляемыми (симптом «рука-лопата»). У больных нарушаются движения при письме, возможность правильного воспроизведения различных поз руки (апраксия позы); они не могут показать без предмета, как совершается то или иное действие (например, как наливают чай в стакан, как закуривают сигарету и т. п.). При сохранности внешней пространственной организации движений нарушается внутренняя проприоцептивная кинестетическая афферентация двигательного акта. При усилении зрительного контроля движения можно в определенной степени скомпенсировать. При поражении левого полушария кинестетическая апраксия обычно носит двухсторонний характер, при поражении правого полушария — чаще проявляется только в одной левой руке.
Вторая форма апраксий, — пространственная апраксия, или апрактоагнозия, — возникает при поражении теменно-затылочных отделов коры на границе 19-го и 39-го полей. Основой данной формы апраксий является расстройство зрительно-пространственных синтезов, нарушение пространственных представлений («верх-низ», «правое-левое» и т. д.). Таким образом, в этих случаях страдает зрительно-пространственная афферентация движений. Пространственная апраксия чаще наблюдается в сочетании со зрительной оптико-пространственной агнозией. Тогда возникает комплексная картина апрактоагнозии. Во всех случаях у больных наблюдаются апраксия позы, трудности выполнения пространственно-ориентированных движений (например, больные не могут застелить постель, одеться и т. п.). Усиление зрительного контроля за движениями им не помогает.
Третья форма апраксий — кинетическая апраксия — связана с поражением нижних отделов премоторной области коры больших полушарий (б-го, 8-го полей — передних отделов «коркового» ядра двигательного анализатора). Она протекает на фоне нарушения автоматизации (временной организации) различных психических функций. Проявляется в виде распада «кинетических мелодий», т. е. нарушения последовательности движений, временной организации двигательных актов. Для этой формы апраксии характерны двигательные персеверации, проявляющиеся в бесконтрольном продолжении раз начавшегося движения.
Кинетическая апраксия проявляется в нарушении самых различных двигательных актов: предметных действий, рисования, письма, — в трудности выполнения графических проб, особенно при серийной организации движений (динамическая апраксия). При поражении нижне-премоторных отделов коры левого полушария (у правшей) кинетическая апраксия наблюдается, как правило, в обеих руках.
Четвертая форма апраксии — регуляторная или префронтальная апраксия — возникает при поражении конвекситальной префронтальной коры кпереди от премоторных отделов; протекает на фоне почти полной сохранности тонуса и мышечной силы.
Она проявляется в виде нарушений программирования движений, отключения сознательного контроля за их выполнением, замены нужных движений моторными шаблонами и стереотипами. При грубом распаде произвольной регуляции движений у больных наблюдаются симптомы эхопраксии в виде бесконтрольных подражательных повторений движений экспериментатора.
Для регуляторной апраксии характерны системные персеверации, т. е. персеверации всей двигательной программы в целом, а не ее отдельных элементов. Такие больные после письма под диктовку на предложение нарисовать треугольник обводят контур треугольника движениями, характерными для письма, и т. п. Наибольшие трудности у этих больных вызывает смена программ движений и действий. В основе этого дефекта лежит нарушение произвольного контроля за осуществлением движения, нарушение речевой регуляции двигательных актов.
Классификация апраксии, созданная А. Р. Лурия, основана главным образом на анализе нарушений двигательных функций у больных с поражением левого полушария головного мозга. В меньшей степени исследованы формы нарушения произвольных движений и действий при поражении различных корковых зон правого полушария; это — одна из актуальных задач современной нейропсихологии.
Вестибулярный тракт
Вестибулярный афферентный путь начинается от рецепторов полукружного канала внутреннего уха, маточки и рецепторов, входящих в состав этого органа. Данный тракт в центральной нервной системе отвечает координацию движений и поддержку равновесия при физических и вестибулярных нагрузках.
Афферентные центростремительные проводящие пути и особенность их строения свидетельствуют о том, человеку нужно прилагать массу усилий, чтобы сохранить здоровье и целостность каждого органа по отдельности и вместе взятых. Каждая составляющая данного пути обеспечивает организм всей необходимой информацией, помогает сразу же обрабатывать ее и приводить в исполнение осуществление всех жизненно важных процессов. Это важно в работе всего организма в целом и отдельных органов.