Article on the topic “Features of the structure and function of subcortical structures.”
- Features of the structure and function of subcortical structures.
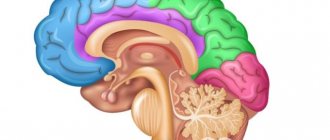
Subcortical brain structures are sections of the brain located between the cerebral cortex and the medulla oblongata. They have an activating effect on the cortex, participate in the formation of all human behavioral reactions, and in maintaining muscle tone.
Subcortical formations include structures located between the cerebral cortex and the medulla oblongata: the thalamus, hypothalamus, basal ganglia, a complex of formations united in the limbic system of the brain, as well as the reticular formation of the brain stem and thalamus. Any afferent excitation that arises from stimulation of receptors in the periphery is transformed at the level of the brain stem into two streams of excitations. One flow along specific paths reaches the projection area of the cortex specific for a given stimulation; the other, through collaterals, enters the reticular formation, from where, in the form of an ascending flow of excitations, it is directed to the cerebral cortex, activating it. The reticular formation has close functional and anatomical connections with the hypothalamus, thalamus, medulla oblongata, limbic system, cerebellum, therefore many types of body activities (breathing, food and pain reactions, motor acts, etc.) are carried out with its mandatory participation.
Afferent flows of excitations from peripheral receptors on the way to the cerebral cortex have numerous synaptic switches in the thalamus. From the lateral group of thalamic nuclei (specific nuclei), excitations are directed along two paths: to the subcortical ganglia and to specific projection zones of the cerebral cortex. The medial group of thalamic nuclei (nonspecific nuclei) serves as a switching point for ascending activating influences that are directed from the stem reticular formation to the cerebral cortex. Close functional relationships between the specific and nonspecific nuclei of the thalamus provide the primary analysis and synthesis of all afferent excitations entering the brain. At low stages of phylogenetic development, the thalamus and limbic formations play the role of the highest center for the integration of behavior, providing all the necessary motor acts aimed at preserving its life. In humans, the highest center of integration is the cerebral cortex.
The limbic system includes a complex of brain structures that plays a leading role in the formation of the main innate human reactions: food, sexual and defensive. It includes the lumbar gyrus, hippocampus, piriformis gyrus, olfactory tubercle, amygdala complex and septal area. The central place among the formations of the limbic system is given to the hippocampus. The hippocampal circle is anatomically established (hippocampus - fornix - mammillary bodies - anterior nuclei of the thalamus - cingulate gyrus - hippocampus), which, together with the hypothalamus, plays a leading role in the formation of emotions. The regulatory influences of the limbic system widely extend to autonomic functions (maintaining a constant internal environment of the body, regulation of blood pressure, respiration, vascular tone, gastrointestinal motility, sexual functions).
The cerebral cortex has constant descending (inhibitory and facilitating) influences on subcortical structures. There are various forms of cyclic interaction between the cortex and subcortical structures, expressed in the circulation of excitations between them. The most pronounced closed cyclic connection exists between the thalamus and the somatosensory area of the cerebral cortex, which functionally constitute a single whole. Cortical-subcortical circulation of excitations can serve as the basis for the formation of conditioned reflex activity of the body.
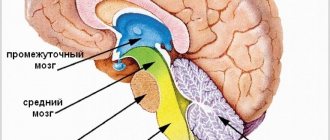
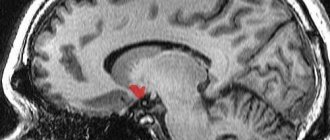
The thalamus (thalamus, visual thalamus) is a structure in which the processing and integration of almost all signals going to the cerebral cortex from the spinal cord, midbrain, cerebellum, and basal ganglia of the brain occurs.
Morphofunctional organization. In the nuclei of the thalamus, information coming from extero-, proprioceptors and interoceptors is switched and thalamocortical pathways begin.
Considering that the geniculate bodies of the thalamus are the subcortical centers of vision and hearing, and the frenulum node and the anterior visual nucleus are involved in the analysis of olfactory signals, it can be argued that the visual thalamus as a whole is a subcortical “station” for all types of sensitivity. Here, irritations from the external and internal environment are integrated and then enter the cerebral cortex.
The visual thalamus is the center of organization and implementation of instincts, drives, and emotions. The ability to receive information about the state of many body systems allows the thalamus to participate in the regulation and determination of the functional state of the body as a whole (this is confirmed by the presence of about 120 multifunctional nuclei in the thalamus). The nuclei form unique complexes that can be divided based on their projection into the cortex into 3 groups: the anterior one projects the axons of its neurons into the cingulate gyrus of the cerebral cortex; medial - in the frontal lobe of the cortex; lateral - into the parietal, temporal, occipital lobes of the cortex. The function of the nuclei is also determined from the projections. This division is not absolute, since one part of the fibers from the thalamic nuclei goes to strictly limited cortical formations, the other to different areas of the cerebral cortex.
The nuclei of the thalamus are functionally divided into specific, nonspecific and associative according to the nature of the pathways entering and exiting them.
Specific nuclei include the anterior ventral, medial, ventrolateral, postlateral, postmedial, lateral and medial geniculate bodies. The latter belong to the subcortical centers of vision and hearing, respectively.
The main functional unit of specific thalamic nuclei are “relay” neurons, which have few dendrites and a long axon; their function is to switch information going to the cerebral cortex from skin, muscle and other receptors.
From specific nuclei, information about the nature of sensory stimuli comes to strictly defined areas of the III-IV layers of the cerebral cortex (somatotopic localization). Dysfunction of specific nuclei leads to loss of specific types of sensitivity, since the nuclei of the thalamus, like the cerebral cortex, have a somatotopic localization. Individual neurons of specific thalamic nuclei are excited by receptors only of their own type. Signals from receptors in the skin, eyes, ear, and muscular system go to specific nuclei of the thalamus. Signals from the interoceptors of the projection zones of the vagus and celiac nerves and the hypothalamus also converge here.
The lateral geniculate body has direct efferent connections with the occipital lobe of the cerebral cortex and afferent connections with the retina and the anterior colliculus. Neurons of the lateral geniculate bodies react differently to color stimulation, turning on and off the light, i.e., they can perform a detector function.
The medial geniculate body (MCC) receives afferent impulses from the lateral lemniscus and from the inferior colliculi. Efferent pathways from the medial geniculate bodies go to the temporal zone of the cerebral cortex, reaching there the primary auditory area of the cortex. MCT has a clear tonotopic pattern. Consequently, already at the level of the thalamus, the spatial distribution of sensitivity of all sensory systems of the body is ensured, including sensory messages from interoreceptors of blood vessels, abdominal organs, and thoracic cavities.
The association nuclei of the thalamus are represented by the anterior mediodorsal, lateral dorsal nuclei and the cushion. The anterior nucleus is connected with the limbic cortex (cingulate gyrus), the mediodorsal - with the frontal lobe of the cortex, the lateral dorsal - with the parietal, the pillow - with the associative zones of the parietal and temporal lobes of the cerebral cortex.
The main cellular structures of these nuclei are multipolar, bipolar triprocess neurons, i.e. neurons capable of performing polysensory functions. A number of neurons change activity only with simultaneous complex stimulation. On polysensory neurons, excitations of different modalities converge, an integrated signal is formed, which is then transmitted to the associative cortex of the brain. Pillow neurons are connected mainly with the associative zones of the parietal and temporal lobes of the cerebral cortex, neurons of the lateral nucleus - with the parietal nucleus, neurons of the medial nucleus - with the frontal lobe of the cerebral cortex.
Nonspecific nuclei of the thalamus are represented by the median center, paracentral nucleus, central medial and lateral, submedial, ventral anterior, parafascicular complexes, reticular nucleus, periventricular and central gray mass. The neurons of these nuclei form their connections according to the reticular type. Their axons rise into the cerebral cortex and contact all its layers, forming not local, but diffuse connections. Nonspecific nuclei receive connections from the RF of the brainstem, hypothalamus, limbic system, basal ganglia, and specific nuclei of the thalamus.
Excitation of nonspecific nuclei causes the generation of specific spindle-shaped electrical activity in the cortex, indicating the development of a sleepy state. Dysfunction of the nonspecific nuclei makes it difficult to develop spindle-shaped activity, that is, the development of a sleepy state.
The brain participates in the formation of the adaptive behavior of an animal, which is based on the principle of a conditioned reflex, as a single integral system. Any stimuli - both conditioned and unconditioned - converge to the same neuron of various subcortical formations, as well as to one neuron of different areas of the cerebral cortex. Studying the mechanisms of interaction between the cortex and subcortical formations in the process of forming the body’s behavioral response is one of the main tasks of modern brain physiology. The cerebral cortex, being the highest authority for the synthesis of afferent excitations, organizes internal nerve connections to perform a reflex act. The reticular formation and other subcortical structures, exerting multiple ascending influences on the cerebral cortex, create only the necessary conditions for the organization of more advanced cortical temporary connections, and as a result, for the formation of an adequate behavioral response of the body. The cerebral cortex, in turn, exerts constant descending (inhibitory and facilitating) influences on subcortical structures. This close functional interaction between the cortex and underlying brain structures lies the basis for the integrative activity of the brain as a whole.
The basal ganglia develop faster than the visual thalamus. Myelination of the structures of the basal ganglia begins in the embryonic period and ends by the first year of life. The motor activity of a newborn depends on the functioning of the globus pallidus. Impulses from it cause general uncoordinated movements of the head, torso, and limbs. In the newborn, the basal ganglia are connected to the optic thalamus, hypothalamus and substantia nigra. With the development of the striatum, the child develops facial movements, and then the ability to sit and stand. At 10 months the child can stand freely. As the basal ganglia and cerebral cortex develop, movements become more coordinated. By the end of preschool age, a balance of cortical-subcortical motor mechanisms is established.
Anatomy of the human subcortex - information:
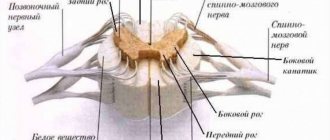
In addition to the gray cortex on the surface of the hemisphere, there are also accumulations of gray matter in its thickness, called the basal ganglia and constituting what is called the subcortex . Unlike the cortex, which has the structure of screen centers, the subcortical nuclei have the structure of nuclear centers.
There are three clusters of subcortical nuclei : corpus striatum, claustrum and corpus amygdaloideum.
1. Coprus striatum, striatum , consists of two parts that are not completely separated from each other - the nucleus caudatus and the nucleus lentiformis.
A. Nucleus caudatus, caudate nucleus , lies above and medial to the nucleus lentiformis, separated from the latter by a layer of white matter called the internal capsule, capsula interna. The thickened anterior part of the caudate nucleus, its head, caput nuclei caudati, forms the lateral wall of the anterior horn of the lateral ventricle, while the posterior thinned section of the caudate nucleus, corpus et cauda nuclei caudati, stretches back along the bottom of the central part of the lateral ventricle; The cauda wraps around the upper wall of the lower horn. On the medial side, the nucleus caudatus is adjacent to the thalamus, separated from it by a strip of white matter, stria terminalis. In front and below, the head of the caudate nucleus reaches the substantia perforata anterior, where it connects with the nucleus lentiformis (with the part of the latter called putamen). In addition to this wide connection of both nuclei on the ventral side, there are also thin strips of gray matter, interspersed with white bundles of the internal capsule. They gave rise to the name “striatum”, corpus striatum.
B. Nucleus lentiformis, lentiform nucleus , lies lateral to the nucleus caudatus and thalamus, separated from them by the capsula interna. On a horizontal section of the hemisphere, the medial surface of the lenticular nucleus, facing the internal capsule, has the shape of an angle with the apex directed towards the middle; the anterior side of the angle is parallel to the caudate nucleus, and the posterior side is parallel to the thalamus. The lateral surface is slightly convex and faces the lateral side of the hemisphere in the region of the insula. Anteriorly and ventrally, as already indicated, the lenticular nucleus merges with the head of the nucleus caudatus. In a frontal section, the lentiform nucleus has the shape of a wedge, the apex of which faces the medial side and the base faces the lateral side.
The lenticular nucleus is divided into three segments by two parallel white layers, laminae medullares, of which the lateral, dark gray, is called the putamen, and the two medial, lighter ones, are collectively called the globus pallidus , globus pallidus. Differing already in its macroscopic appearance, the globus pallidus also has a histological structure that is different from other parts of the striatum.
Phylogenetically, globus pallidus is an older formation (paleostriatum) than putamen and nucleus caudatus (neostriatum). In view of all these features, globus pallidus is currently classified into a special morphological unit called pallidum, while the designation striatum is reserved only for putamen and nucleus caudatus. As a result, the term “lenticular nucleus” loses its previous meaning and can only be used in a purely topographical sense, and instead of the previous name corpus striatum, the caudate and lenticular nucleus are called the striopallidal system.
The striopallidal system is the main part of the extrapyramidal system, and in addition, it is the highest regulatory center of autonomic functions in relation to heat regulation and carbohydrate metabolism, dominating over similar autonomic centers in the hypothalamus.
2. Claustrum, fence , is a thin plate of gray matter located in the region of the insula, between it and the putamen. It is separated from the latter by a layer of white matter, capsula externa, and from the insular cortex by a layer called capsula extrema.
3. Corpus amygdaloideum, the amygdala, is located under the putamen at the anterior end of the temporal lobe. Corpus amygdaloideum apparently belongs to the subcortical olfactory centers and to the limbic system. It ends in a bundle of fibers coming from the olfactory lobe and substantia perforata anterior, noted in the description of the thalamus under the name stria terminalis.
The limbic system is a complex of formations of the telencephalon, diencephalon and mesencephalon, involved in the regulation of various autonomic functions, maintaining the constancy of the internal environment of the body (homeostasis) and in the formation of emotionally charged behavioral reactions. Therefore, some authors designate the limbic system as the “visceral brain.” The main part of it consists of the structures of the cerebral cortex, located mainly on the medial surface of its hemispheres, and subcortical formations closely associated with them, namely: amygdaloid region, stria terminalis, hypothalamus, hippocampus, fornix, septal region, mammillary bodies, mastoid-thalamic fascicle , thalamus, cingulate gyrus. On the medial surface of the cerebral hemispheres, the limbic system is represented by the cingulate and parahippocampal gyri.
T.V. Melnikova
Nizhny Novgorod, Russia
Studying the role of subcortical structures and cortical-subcortical interaction in the organization of higher mental functions (HMF) remains an urgent problem.
The subcortical formations that are part of the activation “energy” block at the same time have their own specific clinical characteristics, and it can be assumed that they are differentially involved in the implementation of cognitive functions.
At the same time, neuropsychological studies indicate heterogeneity of activating systems, manifested by a decrease in motor, speech, and intellectual activity when the left hemisphere is damaged, and features of the emotional and personal sphere when the right hemisphere is damaged.
Impairments in cognitive functions with subcortical lesions depend on the prevalence of the pathological process and its etiology (vascular or tumor diseases of the brain).
Data on the specific participation of the subcortical nuclei in mental activity, the formation of interhemispheric asymmetry at the subcortical level were obtained when studying HMF in patients with parkinsonism before and after stereotactic destruction (Korsakova, Moskovichiute 1985).
The role of subcortical structures in the formation of cognitive impairment is analyzed in progressive dementia (Korsakova et al., 1991), Parkinson’s disease (Zakharov et al., 1999), and arterial-venous deep structures of the brain (Buklina, 1999).
The predominance of dysregulatory disorders (with lacunar strokes of the visual thalamus) was revealed when comparing disorders of higher mental functions and MRI (Yakhno et al., 2002).
The visual thalamus is considered a strategically important area for the implementation of cognitive functions, so the study of neuropsychological deficits in lesions of its various parts is of interest to researchers.
A small number of publications are devoted to neuropsychological research in limited thalamic strokes in the domestic and foreign literature. All studies show a tendency to associate the occurrence of cognitive impairment with damage to certain vascular beds and nuclei of the thalamus (Kalashnikova et al., 1998, Yakhno et al., 2002).
In the acute period, as well as with bilateral paramedian lesions, a decrease in the level of wakefulness, stupor, memory impairment such as Korsakoff syndrome, and hallucinatory syndrome are noted. Clinical manifestations observed in adolescence are considered to be a consequence of neurodynamic disorders.
With lacunar lesions of the visual thalamus in the left hemisphere, disturbances in speech function (poor articulation, slowness of speech, paraphasia), intellectual, visuospatial, and mnestic disturbances are noted.
The complex function of the visual thalamus is due to the presence of specific and nonspecific, secondary and tertiary groups of nuclei, as well as numerous connections: somatosensory and reticulotamic pathways, connections with the orbital and prefrontal, parietal and occipital cortex, cingulate gyrus, mammillary bodies. Typical (posterolateral) thalamic Jerin-Roussy syndrome, resulting from occlusion of the thalamo-genicular artery, is characterized by transient contralateral hemiparesis, contralateral hemianesthesia for tactile and especially deep sensation, hemihypesthesia for pain and temperature sensitivity, spontaneous pain in the involved areas, hemiataxia, astereognosis , contralateral choreoathetosis.
Anterolateral thalamic syndrome is associated with blockage of the thalamic perforating artery and includes intention tremor, choreoathetosis, and thalamic brush.
Comparison of the results of using a highly sensitive neuropsychological method and CT data for lacunar strokes of the visual thalamus allows us to assess the main components of the formation of the neuropsychological syndrome in this pathology and the possibility of lateralization of functions at this level.
Lacunar thalamic strokes according to computed tomography were detected in 7 patients (5 women and 2 men). The average age of those examined was 57 years.
In three cases, computed tomography of the brain indicated the localization of the process in the thalamus cushion, in the lateral and middle sections; in 4 cases, the localization of the lesion within the visual thalamus was not precisely determined. Right hemisphere localization was established in 5 patients, left hemisphere in 2.
The examination was carried out 1-3 weeks after acute stroke. In no case were there any severe memory impairments, disorientation in place and time, or hallucinatory syndrome. Only one patient had Dejerine-Roussy syndrome, with symptoms of hemihypesthesia, ataxia, and burning pain in the contralateral half of the body. In the rest, focal symptoms were mild and manifested as mild transient hemiparesis, intermittent sensory disturbances, and ataxia.
asymmetry of the hemisphere reaction – previous | next – thalamus
A. R. Luria and psychology of the 21st century. Content