Статья на тему «Особенности строения и функции подкорковых структур.»
- Особенности строения и функции подкорковых структур.
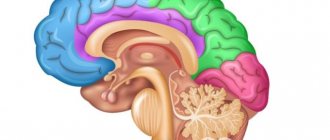
Подкорковые структуры мозга — отделы головного мозга, расположенные между корой больших полушарий и продолговатым мозгом. Оказывают активирующее влияние на кору, участвуют в формировании всех поведенческих реакций человека, в поддержании мышечного тонуса.
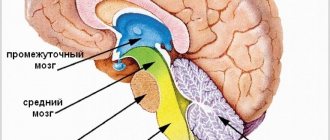
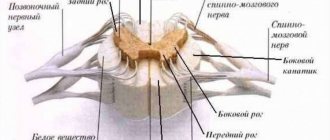
К подкорковым образованиям относят структуры, находящиеся между корой головного мозга и продолговатым мозгом: таламус, гипоталамус, базальные ядра, комплекс образований, объединяемых в лимбическую систему мозга, а также ретикулярную формацию ствола мозга и таламуса. Любое афферентное возбуждение, возникшее при раздражении рецепторов на периферии, на уровне ствола мозга трансформируется в два потока возбуждений. Один поток по специфическим путям достигает специфической для данного раздражения проекционной области коры; другой — по коллатералям попадает в ретикулярную формацию, откуда в виде восходящего потока возбуждений направляется к коре больших полушарий, активируя ее. Ретикулярная формация имеет тесные функциональные и анатомические связи с гипоталамусом, таламусом, продолговатым мозгом, лимбической системой, мозжечком, поэтому многие виды деятельности организма (дыхание, пищевая и болевая реакции, двигательные акты и др.) осуществляются при ее обязательном участии.
Афферентные потоки возбуждений от периферических рецепторов на пути к коре больших полушарий имеют многочисленные синаптические переключения в таламусе. От латеральной группы ядер таламуса (специфические ядра) возбуждения направляются по двум путям: к подкорковым ганглиям и к специфическим проекционным зонам коры мозга. Медиальная группа ядер таламуса (неспецифические ядра) служит местом переключения восходящих активирующих влияний, которые направляются от стволовой ретикулярной формации в кору мозга. Тесные функциональные взаимосвязи между специфическими и неспецифическими ядрами таламуса обеспечивают первичный анализ и синтез всех афферентных возбуждений, поступающих в головной мозг. На низких ступенях филогенетического развития, таламус и лимбические образования играют роль высшего центра интеграции поведения, обеспечивая все необходимые двигательные акты, направленные на сохранение его жизни. У человека высшим центром интеграции является кора больших полушарий.
К лимбической системе относят комплекс структур головного мозга, который играет ведущую роль в формировании основных врожденных реакций человека: пищевых, половых и оборонительных. Он включает в себя поясничную извилину, гиппокамп, грушевидную извилину, обонятельный бугорок, миндалевидный комплекс и область перегородки. Центральное место среди образований лимбической системы отводится гиппокампу. Анатомически установлен гиппокампальный круг (гиппокамп -свод мозга- мамиллярные тела -передние ядра таламуса -поясная извилина -гиппокамп), который вместе с гипоталамусом играет ведущую роль в формировании эмоций. Регуляторные влияния лимбической системы широко распространяются на вегетативные функции (поддержание постоянства внутренней среды организма, регуляция кровяного давления, дыхания, тонуса сосудов, моторики желудочно – кишечного тракта, половых функций).
Кора больших полушарий оказывает постоянные нисходящие (тормозные и облегчающие) влияния на подкорковые структуры. Существуют различные формы циклического взаимодействия между корой и подкорковыми структурами, выражающиеся в циркуляции возбуждений между ними. Наиболее выраженная замкнутая циклическая связь существует между таламусом и соматосенсорной областью коры мозга, составляющими в функциональном отношении единое целое. Корково-подкорковая циркуляция возбуждений может служить основой для формирования условно — рефлекторной деятельности организма.
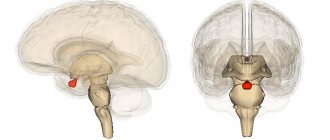
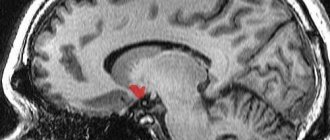
Таламус (thalamus, зрительный бугор) — структура, в которой происходит обработка и интеграция практически всех сигналов, идущих в кору большого мозга от спинного, среднего мозга, мозжечка, базальных ганглиев головного мозга.
Морфофункциональная организация. В ядрах таламуса происходит переключение информации, поступающей от экстеро-, проприорецепторов и интероцепторов и начинаются таламокортикальные пути.
Учитывая, что коленчатые тела таламуса являются подкорковыми центрами зрения и слуха, а узел уздечки и переднее зрительное ядро участвуют в анализе обонятельных сигналов, можно утверждать, что зрительный бугор в целом является подкорковой «станцией» для всех видов чувствительности. Здесь раздражения внешней и внутренней среды интегрируются, после чего поступают в кору большого мозга.
Зрительный бугор является центром организации и реализации инстинктов, влечений, эмоций. Возможность получать информацию о состоянии множества систем организма позволяет таламусу участвовать в регуляции и определении функционального состояния организма в целом (подтверждением тому служит наличие в таламусе около 120 разнофункциональных ядер). Ядра образуют своеобразные комплексы, которые можно разделить по признаку проекции в кору на 3 группы: передняя проецирует аксоны своих нейронов в поясную извилину коры большого мозга; медиальная — в лобную долю коры; латеральная — в теменную, височную, затылочную доли коры. По проекциям определяется и функция ядер. Такое деление не абсолютно, так как одна часть волокон от ядер таламуса идет в строго ограниченные корковые образования, другая — в разные области коры большого мозга.
Ядра таламуса функционально по характеру входящих и выходящих из них путей делятся на специфические, неспецифические и ассоциативные.
К специфическим ядрам относятся переднее вентральное, медиальное, вентролатеральиое, постлатеральное, постмедиальное, латеральное и медиальное коленчатые тела. Последние относятся к подкорковым центрам зрения и слуха соответственно.
Основной функциональной единицей специфических таламических ядер являются «релейные» нейроны, у которых мало дендритов и длинный аксон; их функция заключается в переключении информации, идущей в кору большого мозга от кожных, мышечных и других рецепторов.
От специфических ядер информация о характере сенсорных стимулов поступает в строго определенные участки III—IV слоев коры большого мозга (соматотопическая локализация). Нарушение функции специфических ядер приводит к выпадению конкретных видов чувствительности, так как ядра таламуса, как и кора большого мозга, имеют соматотопическую локализацию. Отдельные нейроны специфических ядер таламуса возбуждаются рецепторами только своего типа. К специфическим ядрам таламуса идут сигналы от рецепторов кожи, глаз, уха, мышечной системы. Сюда же конвергируют сигналы от интерорецепторов зон проекции блуждающего и чревного нервов, гипоталамуса.
Латеральное коленчатое тело имеет прямые эфферентные связи с затылочной долей коры большого мозга и афферентные связи с сетчаткой глаза и с передними буграми четверохолмий. Нейроны латеральных коленчатых тел по-разному реагируют на цветовые раздражения, включение, выключение света, т. е. могут выполнять детекторную функцию.
В медиальное коленчатое тело (МТК) поступают афферентные импульсы из латеральной петли и от нижних бугров четверохолмий. Эфферетные пути от медиальных коленчатых тел идут в височную зону коры большого мозга, достигая там первичной слуховой области коры. МКТ имеет четкую тонотопичность. Следовательно, уже на уровне таламуса обеспечивается пространственное распределение чувствительности всех сенсорных систем организма, в том числе сенсорных посылок от интерорецепторов сосудов, органов брюшной, грудной полостей.
Ассоциативные ядра таламуса представлены передним медиодорсальным, латеральным дорсальным ядрами и подушкой. Переднее ядро связано с лимбической корой (поясной извилиной), медиодорсальное — с лобной долей коры, латеральное дорсальное — с теменной, подушка — с ассоциативными зонами теменной и височной долями коры большого мозга.
Основными клеточными структурами этих ядер являются мультиполярные, биполярные трехотростчатые нейроны, т. е. нейроны, способные выполнять полисенсорные функции. Ряд нейронов изменяет активность только при одновременном комплексном раздражении. На полисенсорных нейронах происходит конвергенция возбуждений разных модальностей, формируется интегрированный сигнал, который затем передается в ассоциативную кору мозга. Нейроны подушки связаны главным образом с ассоциативными зонами теменной и височной долей коры большого мозга, нейроны латерального ядра — с теменной, нейроны медиального ядра — с лобной долей коры большого мозга.
Неспецифические ядра таламуса представлены срединным центром, парацентральным ядром, центральным медиальным и латеральным, субмедиальным, вентральным передним, парафасцикулярным комплексами, ретикулярным ядром, перивентрикулярной и центральной серой массой. Нейроны этих ядер образуют свои связи по ретикулярному типу. Их аксоны поднимаются в кору большого мозга и контактируют со всеми ее слоями, образуя не локальные, а диффузные связи. К неспецифическим ядрам поступают связи из РФ ствола мозга, гипоталамуса, лимбической системы, базальных ганглиев, специфических ядер таламуса.
Возбуждение неспецифических ядер вызывает генерацию в коре специфической веретенообразной электрической активности, свидетельствующей о развитии сонного состояния. Нарушение функции неспецифических ядер затрудняет появление веретенообразной активности, то есть развитие сонного состояния.
Мозг участвует в формировании приспособительного поведения животного, в основе которого лежит принцип условного рефлекса, как единая целостная система. Любые раздражители — как условные, так и безусловные — конвергируют к одному и тому же нейрону различных подкорковых образований, а также к одному нейрону различных областей коры больших полушарий. Изучение механизмов взаимодействия коры и подкорковых образований в процессе формирования поведенческой реакции организма — одна из основных задач современной физиологии головного мозга. Кора больших полушарий, являясь высшей инстанцией синтеза афферентных возбуждений, организует внутренние нервные связи для совершения ответного рефлекторного акта. Ретикулярная формация и другие подкорковые структуры, оказывая множественные восходящие влияния на кору головного мозга, создают лишь необходимые условия для организации более совершенных корковых временных связей, а в результате этого — и для формирования адекватной поведенческой реакции организма. Кора больших полушарий в свою очередь оказывает постоянные нисходящие (тормозные и облегчающие) влияния на подкорковые структуры. В этом тесном функциональном взаимодействии между корой и нижележащими образованиями головного мозга заключена основа интегративной деятельности мозга как единого целого.
Базальные ядра развиваются быстрее, чем зрительные бугры. Миелинизация структур базальных ядер начинается еще в эмбриональном периоде, а заканчивается к первому году жизни. Двигательная активность новорожденного зависит от функционирования бледного шара. Импульсы от него вызывают общие нескоординированные движения головы, туловища, конечностей. У новрожденного базальные ядра связаны со зрительными буграми, гипоталамусом и черной субстанцией. При развитии полосатого тела у ребенка появляются мимические движения, а затем умения сидеть и стоять. В 10 месяцев ребенок может свободно стоять. По мере развития базальных ядер и коры головного мозга движения становятся более координированными. К концу дошкольного возраста устанавливается равновесие корково-подкорковых двигательных механизмов.
Анатомия Подкорки человека — информация:
Кроме серой коры на поверхности полушария, имеются еще скопления серого вещества в его толще, именуемые базальными ядрами и составляющие то, что для краткости называют подкоркой. В отличие от коры, имеющей строение экранных центров, подкорковые ядра имеют строение ядерных центров.
Различают три скопления подкорковых ядер: corpus striatum, claustrum и corpus amygdaloideum.
1. Coprus striatum, полосатое тело, состоит из двух не вполне отделенных друг от друга частей — nucleus caudatus и nucleus lentiformis.
A. Nucleus caudatus, хвостатое ядро, лежит выше и медиальнее nucleus lentiformis, отделяясь от последнего прослойкой белого вещества, называемой внутренней капсулой, capsula interna. Утолщенная передняя часть хвостатого ядра, его головка, caput nuclei caudati, образует латеральную стенку переднего рога бокового желудочка, задний же утонченный отдел хвостатого ядра, corpus et cauda nuclei caudati, тянется назад по дну центральной части бокового желудочка; cauda заворачивается на верхнюю стенку нижнего рога. С медиальной стороны nucleus caudatus прилегает к таламусу, отделяясь от него полоской белого вещества, stria terminalis. Спереди и снизу головка хвостатого ядра доходит до substantia perforata anterior, где она соединяется с nucleus lentiformis (с частью последнего, называемой putamen). Кроме этого широкого соединения обоих ядер с вентральной стороны, имеются еще тонкие полоски серого вещества, располагающиеся вперемешку с белыми пучками внутренней капсулы. Они послужили причиной названия «полосатое тело», corpus striatum.
Б. Nucleus lentiformis, чечевицеобразное ядро, залегает латерально от nucleus caudatus и таламуса, отделенное от них capsula interna. На горизонтальном разрезе полушария медиальная поверхность чечевицеобразного ядра, обращенная к внутренней капсуле, имеет форму угла с верхушкой, направленной к середине; передняя сторона угла параллельна хвостатому ядру, а задняя — таламусу. Латеральная поверхность немного выпукла и обращена к латеральной стороне полушария в области островка. Спереди и вентрально, как было уже указано, чечевицеобразное ядро сливается с головкой nucleus caudatus. На фронтальном разрезе чечевицеобразное ядро имеет форму клина, верхушка которого обращена в медиальную сторону, а основание — в латеральную.
Чечевицеобразное ядро двумя параллельными белыми прослойками, laminae medullares, разделяется на три членика, из которых латеральный, темно-серого цвета, называется скорлупой, putamen, а два медиальных, более светлых, носят вместе название бледного шара, globus pallidus. Отличаясь уже по своему макроскопическому виду, globus pallidus имеет также и гистологическую структуру, отличную от других частей полосатого тела.
Филогенетически globus pallidus представляет более старое образование (paleostriatum), чем putamen и nucleus caudatus (neostriatum). Ввиду всех этих особенностей globus pallidus в настоящее время выделяют в особую морфологическую единицу под названием pallidum, тогда как обозначение striatum оставляют только за putamen и nucleus caudatus. Вследствие этого термин «чечевицеобразное ядро» теряет свое прежнее значение и может употребляться только в чисто топографическом смысле, а вместо прежнего названия corpus striatum хвостатое и чечевицеобразное ядро именуют стриопаллидарной системой.
Стриопаллидарная система представляет собой главную часть экстрапирамидной системы, а кроме того, она является высшим регулирующим центром вегетативных функций в отношении теплорегуляции и углеводного обмена, доминирующим над подобными же вегетативными центрами в hypothalamus.
2. Claustrum, ограда, представляет тонкую пластинку серого вещества, заложенную в области островка, между ним и putamen. От последнего она отделяется прослойкой белого вещества, capsula externa, а от коры островка — прослойкой, носящей название capsula extrema.
3. Corpus amygdaloideum, миндалевидное тело, расположено под putamen в переднем конце височной доли. Corpus amygdaloideum, по-видимому, относится к подкорковым обонятельным центрам и к лимбической системе. В нем оканчивается идущий из обонятельной доли и substantia perforata anterior пучок волокон, отмеченный при описании таламуса под названием stria terminalis.
Лимбическая система представляет комплекс образований конечного, промежуточного и среднего мозга, участвующий в регуляции различных вегетативных функций, поддержании постоянства внутренней среды организма (гомеостаза) и в формировании эмоционально окрашенных поведенческих реакций. Поэтому некоторые авторы обозначают лимбическую систему как «висцеральный мозг». Основную часть ее составляют структуры коры большого мозга, расположенные преимущественно на медиальной поверхности его полушарий, и тесно связанные с ними подкорковые образования, а именно: амигдалоидная область, конечная полоска, гипоталамус, гиппокамп, свод, септальная область, сосцевидные тела, сосцевидно-таламический пучок, таламус, поясная извилина. На медиальной поверхности полушарий большого мозга лимбическая система представлена поясной и парагиппокампальной извилинами.
Т.В. Мельникова
Нижний Новгород, Россия
Изучение роли подкорковых структур и корково-подкоркового взаимодействия в организации высших психических функций (ВПФ) остается актуальной проблемой.
Входящие в состав активационного “энергетического” блока подкорковые образования имеют в то же время свои специфические клинические характеристики, и можно полагать, что они дифференцированно участвуют в реализации когнитивных функций.
Вместе с этим нейропсихологические исследования указывают на неоднородность активирующих систем, проявляющуюся снижением моторной, речевой, интеллектуальной активности при поражении левого полушария, особенностями эмоционально-личностной сферы при поражении правого.
Нарушения когнитивных функций при подкорковых поражениях зависят от распространенности патологического процесса и его этиологии (сосудистые или опухолевые заболевания мозга).
Данные о специфическом участии подкорковых ядер в психической деятельности, формировании межполушарной асимметри на подкорковом уровне получены при изучении ВПФ у больных паркинсонсонизмом до и после стереотаксических деструкций (Корсакова, Московичюте 1985).
Роль подкорковых структур в формировании когнитивных нарушений анализируется при прогрессирующей деменции (Корсакова и др., 1991), при болезни Паркинсона (Захаров и др., 1999), артери-венозных глубинных структурах мозга (Буклина, 1999).
Преобладание дисрегуляторных расстройств (при лакунарных инсультах зрительного бугра) выявлено при сопоставлении нарушений высших психических функций и МРТ (Яхно и др., 2002).
Зрительный бугор считается стратегически важной областью для реализации когнитивных функций, поэтому изучение нейропсихологического дефицита при поражениях различных его отделов представляет интерес для исследователей.
Нейропсихологическим исследованием при ограниченных таламических инсультах в отечественной и зарубежной литературе посвящено небольшое число публикаций. Во всех работах прослеживаются тенденции связывать возникновение когнитивных нарушений с поражением определенных сосудистых бассейнов и ядер зрительного бугра (Калашникова и др., 1998, Яхно и др., 2002).
В остром периоде, а также при двусторонних парамедианных поражениях отмечают снижение уровня бодрствования, сопор, нарушение памяти типа Корсаковского синдрома, галлюцинаторный синдром. Клинические проявления, наблюдаемые в подростковом периоде, рассматриваются как следствие нейродинамических нарушений.
При лакунаторных поражениях зрительного бугра в левом полушарии отмечают нарушения речевой функции (плохую артикуляцию, замедленность речи, парафазии), интеллектуальные, зрительно-пространственные, мнестические нарушения.
Сложная функция зрительного бугра обусловлена наличием специфических и неспецифических, вторичных и третичных групп ядер, а также многочисленными связями: соматосенсорными и ретикулотамическими путями, связями с орбитальной и префронтальной, теменной и затылочной корой, поясной извилиной, маммилярными телами. Типичный (задне-латеральный) таламический синдром Джерина-Русси, возникающий в результате закупорки таламо-коленчатой артерии, характеризуется преходящими контрлатеральным гемипарезом, контрлатеральной гемианестезией для тактильной и особенно глубокой чувствительности, гемигипестезией болевой и температурной чувствительности, спонтанной болью в вовлеченных областях, гемиатаксией, астереогнозом, контрлатеральным хореоатетозом.
Передне-боковой таламический синдром связан с закупоркой таламо-перфорирующей артерии и включает интенционный тремор, хореоатетоз, таламическую кисть.
Сопоставление результатов использования высокочувствительного нейропсихологического метода и данных КТ при лакунарных инсультах зрительного бугра позволяет оценить основные составляющие формирования нейропсихологического синдрома при этой патологии и возможность латерализации функций на этом уровне.
Лакунарные таламические инсульты по данным компьютерной томографии выявлены у 7 больных (5 женщин и 2 мужчин). Среднии возраст обследованных – 57 лет.
В трех случаях при компьютерной томографии головного мозга имелись указания на локализацию процесса в подушке зрительного бугра, в боковых и средних отделах, в 4 случаях локализация очага в пределах зрительного бугра точно не была определена. Правополушарная локализация была установлена у 5 больных, левополушарная у 2.
Обследование проводилось через 1-3 недели после острого инсульта. Ни в одном случае не было отмечено грубых нарушений памяти, дезориентации в месте и времени, галлюцинаторного синдрома. Только у одного больного был выявлен синдром Дежерина-Русси, с явлениями гемигипестезии, атаксии, жгучими болями в контрлатеральной половине тела. У остальных – очаговая симптоматика была легко выражена и проявлялась легкими преходящими гемипарезами, непостоянными нарушениями чувствительности, атаксией.
асимметрия реакции полушарий – предыдущая | следующая – зрительный бугор
А. Р. Лурия и психология XXI века. Содержание