Продолговатый мозг локализуется в заднем отделе головного мозга и связывает его со спинным отделом ЦНС. Сформирован из структур ромбовидного мозга и является частью ствола. Контролирует первостепенные процессы организма – дыхание и кровообращение.
Так как продолговатый мозг связывает церебрум и спинной отдел ЦНС, то совмещает функции и строение обеих структур.
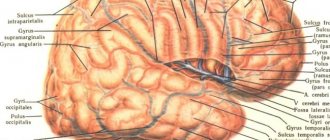
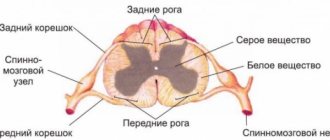
Мозг в разрезе
Строение продолговатого мозга
Продолговатый отдел расположен на стыке спинного и головного отдела ЦНС. Анатомически он находится в области выхода корешков 1-го шейного спинномозгового нерва, а сверху ограничен мостом. По внешнему виду напоминает луковицу. Его высота у взрослого человека составляет примерно 2-3 см.
Несмотря на относительном маленький размер, он выполняет множество функций, без которых особь не смогла бы существовать.
Его структуры состоят из белого и серого вещества, что обуславливает его двойственные функции – он сочетает в себе признаки спинного и конечного мозга с небольшими отличиями: его внутренняя часть состоит из компактно расположенных скоплений серого вещества, а внешняя – из белого. Из задней части продолговатого отдела к спинному тянутся 2 канатика, которые объединяют их.
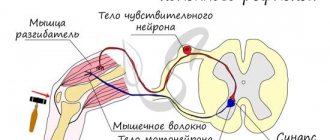
Как известно, большие полушария управляют противоположными сторонами тела: левая часть отвечает правую сторону и, наоборот. Эта особенность центральной нервной системы осуществляется за счет характерного строения продолговатого мозга: он реализует проводниковую функцию, которая состоит в передаче импульсов возбуждения от собственных ядер к нервной ткани других отделов, но не прямым способом, а крест-на-крест.
Размеры и строение продолговатого мозга меняются с возрастом. Например, в отличие от взрослых, у младенцев эта часть ЦНС больше относительно других отделов. Окончательный вид он приобретает к 7 годам.
Границы нахождения
Внешняя структура
В строении продолговатого мозга выделяют 2 части: внутреннюю и внешнюю. Рассмотрим подробнее внешний вид этого отдела и особенности его строения точки зрения анатомии.
Продолговатый мозг представляет собой перевернутый конус: его широкий конец находится сверху, узкий – внизу. В центральной части лицевой стороны располагается передняя срединная щель. Она является продолжением центральной борозды спинного отдела ЦНС.
По сторонам от нее находятся пирамиды, которые переходят в лицевые канаты спинного мозга, состоящие из скопления нервных клеток. На задней стороне продолговатого мозга видна задняя средняя борозда, переходящая в спинной отдел.
Подробное строение
Рядом с ней расположены хвостовые канатики, которые соединены с бороздами спинного отдела ЦНС. Эти канатики являются нижней пограничной чертой продолговатого отдела, а верхней стала для него базальная граница, проходящая в области Варолиева моста.
Внутренняя структура
Внутреннее строение продолговатого мозга эффективнее рассматривать в поперечном разрезе, сделанном на уровне олив. Если смотреть на него с лицевой стороны, то анатомия выглядит следующим образом: от парных возвышений (пирамид) на внешней стороне вглубь структуры отходят тракты преднамеренных моторных импульсов.
Рядом с ними располагаются оливы, охватывающие базальные ядра, корешки подъязычного нерва, идущие к языковым мышцам, аксоны и нервные клетки, сгруппированные в ядра. От каждого ядра к мозжечку проходят длинные волокна, связывающие их между собой. Они образуют оливомозжечковый путь.
Вблизи оливы также располагаются:
- Волокна языкоглоточного нерва;
- Волокна блуждающего нерва;
- Волокна добавочного нерва.
В латеральной (боковой) стороне продолговатого отдела залегают эфферентные проводниковые тракты:
- Руброспинальный;
- Тектоспинальный;
- Ретикулоспинальный;
- Вестибулоспинальный.
На уровне пирамид большая часть аксонов (до 80%), формирующих эфферентные пути, перекрещиваются между собой. Остальные также формируют перекрест и переходят на другую сторону вниз, к спинному отделу ЦНС.
В задней части продолговатого мозга от спинного мозга в верхнележащие отделы ЦНС тянутся 4 проводящие чувствительные тракта:
- Тонкий;
- Клиновидный;
- Спиноталамический;
- Спиномозжечковый.
Они являются естественным продолжением аналогичных структур спинного мозга.
Основные ядра продолговатого мозга
Серое вещество продолговатого мозга обрабатывает данные, прибывающие в ЦНС от всех рецепторов тела человека. Оно представляет собой скопление структур нейронов (тел и частично отростков), компактно расположенное в толще белого вещества.
Выделяют:
- Ядра 9-12 пар черепных нервов;
- Центры кровообращения и дыхания;
- Ядро оливы;
- Ретикулярная формация.
В продолговатом отделе насчитывают 5 пар ядер черепных нервов. Они сконцентрированы в хвостовой части большого мозга пониже дна 4 желудочка:
- Преддверно-улитковый нерв (VIII пара). Состоит из улитковой и преддверной частей. Отвечает за проведение слуховых импульсов, а также сигналов, отходящих от вестибулярного отдела внутреннего уха.
- Языкоглоточные нервы (IX пара). Включает в себя сенсорные, двигательные и секреторные волокна. Моторные аксоны реализуют двигательную функцию – контролируют активность мышц глоточного канала и ротовой полости. С помощью чувствительных волокон к ядру поступает информация, полученная из пищевкусовой сенсорной системы задней части языка. Парасимпатические волокна отвечают за секреторную функцию в ротовой полости: контролируют слюновыделительные процессы.
- Блуждающий нерв (X пара). Состоит из 3 видов ядер: вегетативных, сенсорных и моторных. Вегетативное ядро выполняет функцию связующего звена церебрума и органов головы, шеи, грудной и брюшной полости. От него простилаются в разные стороны входящие и нисходящие волокна. К чувствительному ядру поступает информация от легких, сердца и других важных систем организма. Моторное ядро контролирует сокращение мышц ротовой полости во время глотания. Здесь также располагается обоюдное ядро, аксоны которого выполняют защитную функцию – активируют процессы кашля, чихания, рвоты.
- Добавочный нерв (XI пара). Представлен 2 ядрами: одно залегает в продолговатом мозге на дне ромбовидной ямки, другое – в переднем роге на уровне 1 сегмента позвоночника. Волокна от него входят в большое затылочное отверстие черепа, объединяются с черепными корешками, формируя ствол добавочного нерва. Затем они выводятся наружу из полости черепа через яремное отверстие. Добавочный нерв выполняет контролирующую функцию – отвечает за моторику грудино-ключично-сосцевидной и трапециевидной мышцы.
- Подъязычный нерв (XII пара). Контролирует моторную функцию языка, так как связан с его шилоязычной, подбородной, прямыми и поперечными мышцами. От его работы зависят безусловные рефлексы: глотание, жевание, сосание. Ядра подъязычного нерва руководят моторикой языка в процессе жевания, произнесения речи и других движений рта. Информация от подъязычного нерва направляется в сенсомоторный участок коры больших полушарий посредством клиновидного и нежного ядра.
В нижней хвостовой части продолговатого отдела располагается центр гемодинамики. Его волокна тесно связаны с ядрами V пары нервов. Ученые предполагают, что именно в центре гемодинамики возникают возбуждающие активизирующие сигналы симпатических волокон на сердечно-сосудистую систему.
Основные ядра
Иногда гемодинамический центр угнетается шоковыми состояниями, травмами, инсультами, отравлениями, метаболическими нарушениями.
Ядро оливы. Представляет собой зубчатую пластинку, функции которой определяются ее структурой – она состоит из серого вещества. Изогнуто в виде подковы. Взаимодействует с зубчатыми ядрами мозжечка, то есть являются магистральными структурами равновесия.
Ядра оливы лучше всех из млекопитающих развита у человека, так как человек больше всех из животного мира находится в вертикальном положении и нуждается в хорошем вестибулярном аппарате. Кроме этого ядра оливы помогают ориентироваться в пространстве: в них осуществляется сравнение громкости звуковых волн, проникающих в правое и левое ухо. Анализируя их, человек поворачивается лицом к источнику звука.
В нижней части продолговатого мозга располагается так называемое «синее пятно» — зона ретикулярной формации. Волокна нейронов этой структуры выделяют специфический гормон, который влияет на степень возбудимости нервных клеток.
«Синее пятно» отвечает за чувство тревоги и внутреннее напряжение, а также обеспечивает связь отдела с органами чувств, спинным мозгом и остальными отделами, выполняя функцию регулятора нервной активности.
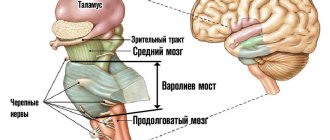
3.4. ПРОДОЛГОВАТЫЙ МОЗГ И МОСТ
3.4.1. Строение продолговатого мозга и моста
Продолговатый мозг и мост (варолиев мост) представляют собой отдел головного мозга, который является непосредственным продолжением спинного мозга. Серое вещество спинного мозга переходит в серое вещество продолговатого мозга и сохраняет черты сегментарного строения. Однако основная часть серого вещества распределена по всему объему заднего мозга в виде обособленных ядер,
разделенных белым веществом.
В продолговатом мозгу симметрично заложены ядра четырех последних пар черепных нервов, осуществляющих афферентную и эфферентную иннервацию головы и внутренних органов. В самой нижней части ромбовидной ямки располагается двигательное ядро подъязычного нерва
(XII пара), иннервирующего мышцы языка.
Двигательное ядро добавочного нерва
(XI пара) является
| Рис. 3.21 Расположение ядер черепных нервов в стволовой части мозга 1 — добавочный нерв, 2 — подъязычный, 3 — преддверно—улитковый, 4 — отводящий, 5 — блоковый, 6 — глазодвигательный, 7 — тройничный, 8 — лицевой, 9 — языкоглоточный, 10 — блуждающий нерв. |
продолжением заднебокового отдела вентрального рога спинного мозга и связано с иннервацией мышц шеи (рис. 3.21).
Довольно поверхностно лежит группа ядер блуждающего нерва
(X пара), который является смешанным т. е. одновременно участвует в вегетативной, двигательной и чувствительной иннервации. Соответственно функции блуждающему нерву принадлежат три ядра. Вегетативное
парасимпатическое ядро блуждающего нерва
дает парасимпатические волокна, идущие к гортани, пищеводу, сердцу, желудку, тонкой кишке, пищеварительным железам (см. разд. 5.3). Чувствительные волокна блуждающего нерва от многих внутренних органов в продолговатом мозгу образуют одиночный путь (солитарныи тракт) и заканчиваются в чувствительном
ядре одиночного пути.
Третье ядро блуждающего нерва — соматическое, двигательное — лежит глубже в латеральном отделе продолговатого мозга и носит название
двойного.
Нейроны этого ядра рефлекторно регулируют последовательность сокращения мышц глотки и гортани при дыхании.
Языкоглоточный нерв
(IX пара), так же как и блуждающий, содержит двигательные, чувствительные и вегетативные волокна. Двигательные волокна участвуют в иннервации мышц ротовой полости и глотки. Чувствительные волокна от вкусовых почек задней трети языка, слизистой оболочки глотки входят вместе с волокнами блуждающего нерва в одиночный путь. Отростки нейронов, формирующих вегетативное ядро языкоглоточного нерва, идут к парасимпатическим ганглиям, которые связаны с иннервацией секреторного аппарата слюнных желез.
На границе продолговатого мозга и моста располагаются ядра преддверно—улиткового,
или
слухового,
нерва (VIII пара). Этот нерв состоит из двух частей:
улитковой и пред дверной. Волокна улитковой части, или собственно слухового нерва, представляют собой аксоны клеток лежащего в улитке спирального узла. Эти волокна оканчиваются в улитковых ядрах
(рис. 3.21).
Волокна преддверной части
формируются аксонами клеток узла преддверия, локализованного на дне внутреннего слухового прохода. Эти волокна заканчиваются в трех вестибулярных ядрах моста:
медиальном
(ядре Швальбе),
латеральном
(ядре Дейтерса) и
верхнем
(ядре Бехтерева). Отростки нейронов второго порядка из этих ядер направляются к ядру шатра мозжечка. Из латерального вестибулярного ядра, которое можно отнести к надсегментарным образованиям, начинается преддверно—спинномозговой путь, идущий в спинной мозг.
В латеральном отделе ретикулярной формации моста расположено ядро лицевого нерва
(VII пара). Этот нерв является смешанным. Афферентные чувствительные волокна лицевого нерва передают информацию от вкусовых рецепторов передней части языка. Вегетативные волокна участвуют в иннервации подчелюстных и подъязычных слюнных желез. Эфферентные волокна лицевого нерва имеют концевые разветвления в мимической мускулатуре лица.
Ядро отводящего нерва
(VI пара) заложено поверхностно в задней половине моста. Моторные волокна этого нерва иннервируют мышцы, двигающие глазное яблоко, чувствительные волокна связаны с проприоцепторами этих мышц.
И наконец, последняя пара черепных нервов, относящаяся к заднему мозгу, — это тройничный нерв
(V пара). Этот нерв смешанный и обладает двигательными и чувствительными ядрами.
Двигательное ядро тройничного нерва
расположено в верхнем отделе покрышки моста. Нейроны этого ядра участвуют в иннервации жевательных мышц, мышцы нёбной занавески и мышцы, напрягающей барабанную перепонку. Длинное, простирающееся через весь мост чувствительное
ядро спинномозгового пути тройничного нерва
содержит клетки, к которым подходят афферентные аксоны от рецепторов кожи лица, слизистой оболочки носа, зубов, надкостницы костей черепа (см. рис. 3.21).
В сером веществе продолговатого мозга расположены ядра чувствительных путей задних столбов спинного мозга. Это тонкое и клиновидное ядра (ядра одноименных пучков),
являющихся путями, которые проводят информацию от тактильных и проприоцепторов, сигнализирующих о положении тела в пространстве. Кроме того, внутри заднего мозга расположено
заднее ядро трапециевидного тела
(верхнее оливное ядро), включенное в восходящий путь, который передает акустическую информацию (см. разд. 4.5.3). По латеральной поверхности продолговатого мозга располагается продолжение боковых столбов спинного мозга.
Центральная часть продолговатого мозга занята ретикулярной,
или сетевидной,
формацией
(веществом), которая представляет собой скопление нервных клеток, напоминающее по принципу организации нервные сети кишечнополостных. Ретикулярные нейроны распределяются и диффузно, и группируются в ядрах, среди которых можно выделить наиболее крупные: гигантоклеточное ретикулярное, мелкоклеточное ретикулярное, латеральное ретикулярное ядро на уровне продолговатого мозга, каудальное, вентрикулярное и оральное ретикулярные ядра — на уровне моста. По цитоархитектоническим особенностям в ретикулярной формации продолговатого мозга выделяют
медиальную часть
с множеством нейронов крупного размера и латеральную, включающую в себя более мелкие элементы. По своей топографии ретикулярная формация ствола мозга является структурой, расположенной на пути всех входных и выходных систем мозга.
К дендритам и соме ретикулярных нейронов подходят коллатерали от аксонов сенсорных восходящих путей, локализованных в центральной и латеральной частях ствола мозга. С другой стороны к нейронам ретикулярной формации подходят коллатерали нисходящих моторных путей, идущих из переднего мозга. От нейронов гигантоклеточного и каудального ретикулярного ядра начинается нисходящий ретикулярно—спинномозговой путь, устанавливающий прямые связи с мотонейронами спинного мозга. Ретикулярная формация имеет связи с ядрами черепных нервов, с мозжечком, промежуточным мозгом и через последний — с корой больших полушарий.
Таким образом, богатство связей ретикулярной формации свидетельствует о том, что она является надсегментарной структурой, приспособленной к регуляции деятельности различных отделов центральной нервной системы.
3.4.2. Рефлексы продолговатого мозга
Продолговатый мозг является жизненно важным отделом нервной системы, где замыкаются дуги целого ряда соматических и вегетативных рефлексов. При участии ядер продолговатого мозга осуществляются цепные рефлексы,
связанные с жеванием и проглатыванием пищи. Так, например, при акте жевания координированная деятельность жевательных мышц, языка, щек, нёба и дна полости рта обеспечивается функцией моторных ядер тройничного и подъязычного нервов. Когда пищевой комок касается нёба, основания языка или задней стенки глотки, возникает цепной рефлекс глотания. От рецепторов задней трети языка, слизистой оболочки глотки возбуждение по чувствительным волокнам языкоглоточного нерва поступает в продолговатый мозг к центру глотательного рефлекса. Из этого центра запрограммированные в нервной системе моторные команды по двигательным волокнам подъязычных, тройничных, языкоглоточных и блуждающих нервов идут к мышцам полости рта, глотки, гортани и пищевода. В результате координированной деятельности этих мышц совершается акт глотания с одновременным перекрытием входов в носоглотку и гортань.
С функцией пищеварительного тракта связаны многие вегетативные рефлексы
заднего мозга. К их числу относится, например рефлекторная регуляция секреции слюнных желез, которая осуществляется с участием ядер лицевого и языкоглоточного нервов, содержащих парасимпатические нейроны. Вегетативное, парасимпатическое ядро блуждающего нерва включено в систему рефлекторной регуляции дыхания, деятельности сердца и тонуса сосудов.
Целая группа рефлексов заднего мозга направлена на поддержание позного тонуса.
В зависимости от источника рецепторных воздействий тонические рефлексы подразделяются на шейные и вестибулярные, или лабиринтные.
Шейные тонические рефлексы
запускаются при возбуждении проприоцепторов мышц шеи. Впервые они описаны голландским физиологом Р. Магнусом на кошках с перерезанным на уровне шатра мозжечка стволом мозга (см. разд. 3.5.2). В чистом виде шейные .рефлексы можно наблюдать при разрушении вестибулярного аппарата, дающего дополнительную информацию о положении головы в пространстве. Как показали опыты Р. Магнуса, запрокидывание головы кошки назад в сагиттальной плоскости вызывает возрастание тонуса мышц—разгибателей передних конечностей и одновременно ослабление экстензорного тонуса задних (рис. 3.22). Напротив, наклон головы вниз вызывает обратный процесс, когда тонус разгибателей передних конечностей снижается, а задних — возрастает. Нарушение равновесия за счет поворота головы вправо или влево относительно продольной оси вызывает компенсаторное усиление тонуса разгибателей тех конечностей, в сторону которых повернута голова. Это так называемый
рефлекс вращения по Магнусу.
Эффекторным звеном шейных рефлексов могут быть не только мышцы конечностей, но и глазная мускулатура. Рефлекторные влияния на глазные мышцы обеспечивают компенсаторную установку глазных яблок при изменении положения головы. Любое отклонение головы вызывает движение глазных яблок в противоположном направлении, и таким образом рефлекторно сохраняется правильная зрительная ориентация животного.
Все шейные рефлексы являются полисинаптическими. Импульсы от проприоцепторов шейных мышц идут в соответствующие центры продолговатого мозга, а оттуда нисходящие команды поступают или к мотонейронам спинного мозга, или к двигательному ядру отводящего нерва и производят определенное влияние на позный тонус и на глазную мускулатуру.
| Рис. 3.22 Позные рефлексы децеребрированного животного А — децеребрационная ригидность; Б, В, Г — шейные тонические рефлексы по Магнусу; объяснение см. в тексте. |
Вестибулярные рефлексы
неразрывно связаны с шейными тоническими рефлексами и в естественных условиях дополняют их. Вестибулярные рефлексы не зависят от положения головы относительно туловища и в чистом виде могут быть получены при фиксации головы по отношению к туловищу или при выключении проприоцепторов шейных мышц новокаиновой блокадой.
Вестибулярные рефлексы, согласно классификации Магнуса, подразделяются на статические и статокинетические. Статические рефлексы
связаны главным образом с возбуждением рецепторов преддверия перепончатого лабиринта и обеспечивают поддержание позы и равновесия тела при самых разнообразных его статических положениях в пространстве. Так, например, если кошку с фиксированной относительно туловища головой поворачивать в пространстве, то в зависимости от положения головы будет наблюдаться различное распределение позного тонуса. В том случае, когда угол между плоскостью горизонта и плоскостью ротовой щели животного составляет 45°, разгибательные мышцы напрягаются максимально. Если же указанный угол равен 135°, наблюдается наименьшее значение тонуса разгибателей.. Между крайними положениями имеется ряд постепенных переходов в степени развития экстензорного тонуса. Это пример тонического вестибулярного рефлекса положения, который осуществляется при участии нейронов латерального вестибулярного ядра (Дейтерса) и идущего от него в спинной мозг преддверно—спинномозгового пути.
К числу статических вестибулярных рефлексов относят рефлексы выпрямления, направленные на переход животного из неестественной позы в обычное для него положение. В качестве наглядного примера можно привести переворачивание кошки в воздухе при падении спиной вниз. За очень короткое время падения животное успевает занять нормальное по отношению к гравитационному полю положение и упасть сразу на все четыре лапы. В процессе переворачивания выпрямительные рефлексы совершаются в определенной последовательности. Сначала за счет вестибулярного выпрямительного рефлекса восстанавливается нормальное положение головы — мордой вниз. Затем изменение положения головы возбуждает проприоцепторы шейных мышц и они запускают шейный выпрямительный рефлекс, в результате которого вслед за головой туловище также возвращается в нормальное положение.
Таким образом, в естественных условиях вестибулярные рефлексы выпрямления дополняются шейными, причем ведущую роль в их осуществлении играет положение головы, где расположены дистантные рецепторы.
Следующая группа вестибулярных рефлексов — статокинетические —
характеризуется тем, что она направлена на поддержание позы при изменении скорости движения животного. Эти рефлексы связаны с возбуждением рецепторов полукружных каналов, которое имеет место при наличии в каналах тока эндолимфы. Обычно направленность статокинетического рефлекса зависит от того, какой полукружный канал раздражается при ускорении. Например, при замедлении поступательного движения возбуждаются рецепторы сагиттального канала и, соответственно, рефлекторные изменения мышечного тонуса будут наклонять туловище вперед, как бы сохраняя равномерное движение.
Ускорение при вращении тела в горизонтальной плоскости возбуждает рецепторы горизонтального полукружного канала и вызывает рефлекторную реакцию глазодвигательного аппарата — горизонтальный нистагм.
Суть этой реакции состоит в том, что в момент ускорения вращения глаза движутся в сторону, противоположную направлению вращения. Затем, достигнув крайнего отклонения, глаза быстро перемещаются обратно в направлении вращения и таким образом в поле зрения попадает другой участок пространства. Нистагм способствует сохранению нормальной зрительной ориентации и обычно используется в диагностических целях для проверки нормального функционирования вестибулярного аппарата. Плоскость нистагма совпадает с плоскостью ускорения, в связи с чем кроме горизонтального нистагма можно встретить вертикальный, диагональный и круговой нистагмы (см. разд. 4.8.4).
К числу статокинетических вестибулярных рефлексов относятся также и так называемые лифтные рефлексы,
которые проявляются в увеличении тонуса мышц разгибателей при линейном ускорении вверх и в повышении тонуса сгибателей при линейном ускорении вниз.
3.4.3. Функции ретикулярной формации стволовой части мозга
В ретикулярной формации продолговатого мозга сосредоточены центры, играющие важную роль в регуляции висцеральных функций.
Это в первую очередь
дыхательный центр,
локализованный в медиальной части ретикулярной формации продолговатого мозга. Еще в прошлом столетии было установлено, что повреждение каудальной части дна IV желудочка приводит к остановке дыхания
(укол Флуранса).
Позднее Н. А. Миславский (1885) выделил в дыхательном центре две функционально различные части:
инспираторную и экспираторную
(или вдыхательную и выдыхательную). Использование методов микростимуляции и микроэлектродной регистрации активности одиночных нейронов позволило уточнить представления о локализации частей дыхательного центра.
Установлено, что инспираторная часть расположена более вентрально, чем экспираторная. Активность одиночных нейронов дыхательного центра может коррелировать с фазами дыхательного цикла. Этот признак позволяет выделить инспираторные нейроны, которые генерируют потенциалы действия в начальную фазу вдоха, и экспираторные нейроны, разряд которых приурочен к фазе выдоха. Наибольшее скопление инспираторных нейронов обнаружено около одиночного (солитарного) пути. В области двойного ядра локализованы инспираторные и экспираторные нейроны. Определенная мозаичность — в расположении дыхательных нейронов свидетельствует о том, что деление дыхательного центра на две части — вдыхательную и выдыхательную — более справедливо в функциональном смысле, чем в анатомическом (см. разд. 9.4.1).
Отличительной чертой нейронов дыхательного центра является способность к автоматизму.
Даже при отсутствии афферентных воздействий активность этих нейронов характеризуется периодичностью, которая определяется спецификой ионных механизмов их клеточной мембраны. Периодичность разрядов дыхательных нейронов может быть обусловлена также наличием взаимных тормозных связей между инспираторными и экспираторными нейронами. Тормозные связи создают реципрокность разрядов дыхательных нейронов, когда появление активности инспираторных нейронов сопровождается торможением разрядов экспираторных и наоборот. В результате реципрокного взаимодействия дыхательных нейронов происходит смена фаз дыхательного цикла.
Вместе с тем смена фаз дыхательного цикла может осуществляться рефлекторно за счет афферентных влияний, которые модулируют дыхательную ритмику. При интенсивном вдохе и растяжении легочной ткани импульсы от механорецепторов легких по афферентным волокнам блуждающего нерва приходят в дыхательный центр и вызывают рефлекторное торможение инспираторных нейронов, одновременно возбуждая экспираторные (рефлекс Геринга—Брейера).
Импульсы от экспираторных нейронов по ретикулярно—спинномозговому пути достигают исполнительных моторных центров спинного мозга и стимулируют начало выдоха.
Периодичность в работе дыхательного центра продолговатого мозга может обеспечиваться также за счет регулирующих влияний со стороны пневмотаксического центра.
Этот центр находится в области моста и также состоит из двух типов нейронов — инспираторных и экспираторных, которые не имеют жесткой приуроченности своих разрядов к фазам дыхательного цикла. Пневмотаксический центр как регулятор периодически затормаживает инспираторную часть дыхательного центра и стимулирует экспираторные нейроны, осуществляя таким образом прекращение вдоха и начало выдоха.
В регуляции функций дыхательного центра могут принимать участие и более высокие этажи головного мозга. Так, например, эмоциональные реакции человека связаны с изменением периодичности в работе дыхательного центра, вызванным импульсацией промежуточного мозга и лимбической коры. Установлено, что электрическое раздражение орбитальной коры может вызвать полную остановку дыхания у человека. Регулирующие воздействия головного мозга обеспечивают произвольную регуляцию дыхания, его коррекцию при разнообразных изменениях жизнедеятельности.
Таким образом, регуляция дыхательной функции
осуществляется иерархической системой нервных центров, расположенных на разных этажах нервной системы и связанных единством действия.
Другим жизненно важным центром ретикулярной формации продолговатого мозга является сосудодвигательный центр,
локализация которого впервые была определена Ф. В. Овсянниковым (1871). Сосудодвигательный, или вазомоторный, центр занимает обширную область продолговатого мозга, простираясь в дорсолатеральном направлении от дна IV желудочка до пирамид (см. разд. 5.5.2). Перерезка спинного мозга на уровне шейных позвонков вызывает у спинальных животных резкое снижение сосудистого тонуса и нарушение регуляторных реакций, связанных с изменением просвета вен и артерий.
Локальная стимуляция ростральных частей сосудодвигательного центра вызывает увеличение сосудистого тонуса, повышение кровяного давления и тахикардию. Стимуляция каудального отдела сосудистого центра, напротив, вызывает расширение сосудов, падение кровяного давления и брадикардию.
Микроэлектродная регистрация электрической активности одиночных нейронов сосудистого центра показала наличие нервных клеток, изменяющих свою фоновую активность синхронно с флуктуациями артериального давления. Среди них можно выделить нейроны, частота разрядов которых возрастает при повышении артериального давления, нейроны, частота фоновой активности которых, напротив, снижается при увеличении артериального давления, и, наконец, нейроны, частота импульсации которых меняется в соответствии с рабочим циклом сердца. Свойства нейронов сосудистого центра менее изучены, чем у нейронов дыхательного центра. Пока неясно, можно ли дифференцировать сосудодвигательные нейроны на «сосудосуживающие» и «сосудорасширяющие». Нерешенным остается также вопрос о наличии тормозного взаимодействия между нейронами сосудодвигательного центра.
Функциональная организация сосудодвигательного центра имеет определенную специфику в плане проекций его нисходящих путей. Эфферентные волокна от его нейронов спускаются в грудной отдел спинного мозга, но заканчиваются там не на мотонейронах, как в случае нисходящих систем дыхательного центра, а на преганглионарных нейронах симпатической нервной системы.
Таким образом, сосудистый тонус регулируется не антагонистическими нисходящими воздействиями, а только одной симпатической сосудосуживающей системой.
Активное состояние этой системы вызывает вазоконстрикцию, а торможение — вазодилатацию (см. разд. 5.4, 8.6). Исключение из этого принципа составляют только некоторые сосуды, имеющие двойную иннервацию — симпатическую и парасимпатическую (сосуды половых органов и слюнных желез).
Рефлекторные влияния на нейроны сосудодвигательного центра осуществляются при возбуждении хемо— и механорецепторов, локализованных в сосудистой стенке. Афферентные волокна от этих рецепторов в составе блуждающего и языкоглоточного нервов достигают продолговатого мозга. Возбуждение механорецепторов дуги аорты, каротидного синуса при повышении артериального давления вызывает торможение активности сосудодвигательного центра и, как следствие, рефлекторное снижение сосудистого тонуса (рефлексы Людвига—Циона, Геринга, Бейнбриджа). Напротив, при повышении давления в системе полых вен наблюдается усиление активности сосудистого центра и вазоконстрикторный эффект. Тонус сосудистого центра может изменяться и При возбуждении хеморецепторов сосудистой стенки, возникающем при изменениях химического состава крови.
Следует отметить, что деятельность сосудодвигательного центра сочетается с функцией моторного ядра блуждающего нерва — двойного ядра, снижающего в норме частоту сердечных сокращений. В связи с этим при вазоконстрикторном эффекте одновременно увеличивается частота сердечных сокращений и, наоборот, при вазодилатации наблюдается замедление сердечной ритмики.
Для ретикулярной формации стволовой части мозга характерны не только вегетативные регулирующие функции, но и участие в нисходящем контроле деятельности двигательных центров спинного мозга.
В 1862 г. И. М. Сеченов установил угнетение спинальных рефлексов при раздражении стволовой части мозга.
Это было открытием центрального торможения и одновременно открытием ретикулоспинальной системы. Однако механизм этого тормозного процесса удалось раскрыть только в 50—е гг. нашего столетия после работ американского нейрофизиолога Г. Мегуна, показавшего, что локальное электрическое раздражение гигантоклеточного ядра ретикулярной формации продолговатого мозга вызывает неспецифическое торможение сгибательных и разгибательных спинальных рефлексов. Эти неспецифические супраспинальные влияния по ретикулярно—спинномозговому пути достигают мотонейронов спинного мозга и увеличивают порог и скрытый период их ответов на рефлекторные воздействия. Дальнейшие исследования показали, что нисходящие влияния ретикулярной формации могут реализовываться не только за счет постсинаптического торможения мотонейронов, но и за счет возникновения длительных тормозных постсинаптических потенциалов в промежуточных нейронах, а также за счет воздействия ретикулоспинальных волокон на терминали афферентных волокон, входящих в спинной мозг.
Таким образом, ослабление рефлекторной деятельности достигается за счет прямого действия на мотонейроны и за счет определенного ослабления сенсорного входа в спинной мозг.
В ходе экспериментов с локальной стимуляцией ретикулярной формации выяснилось наличие зон, дающих эффект противоположной полярности, т. е. облегчающее влияние на спинномозговые рефлексы.
Так, например, электрическое раздражение латеральных зон ретикулярной формации моста снижает порог и укорачивает скрытый период спинальных рефлексов. При стимуляции медиальных ядер ретикулярной формации заднего мозга в мотонейронах мышц—сгибателей кошки регистрируются коротколатентные возбуждающие постсинаптические потенциалы (ВПСП).
Данный факт свидетельствует о наличии (помимо диффузных неспецифических ретикулоспинальных проекций) моносинаптических нисходящих путей специфического действия, которые участвуют в нисходящем контроле деятельности спинного мозга.
Следовательно, ретикулярная формация как один из двигательных центров стволовой части мозга может выступать не только в роли регулятора возбудимости спинальных мотонейронов, но и принимать участие в процессах, связанных с поддержанием позы и организацией целенаправленных движений.
Благодаря работам Г. Мегуна и Дж. Моруцци наряду с неспецифическими нисходящими влияниями ретикулярной формации ствола были открыты ее восходящие, активирующие влияния на кору головного мозга.
Если через хронически вживленные электроды раздражать центральные части ретикулярной формации ствола, то кошка, находящаяся в сонном состоянии, пробуждается и у нее появляется ориентировочная реакция. Эта поведенческая реакция пробуждения сопровождается характерными изменениями частотного спектра электроэнцефалограммы, переходом от регулярных, высоковольтных колебаний α—ритма к низковольтным колебаниям. (β—ритма (см. разд. 3.11.1). Данная электроэнцефалографическая реакция получила название
реакции десинхронизации.
Она имеет генерализованный характер и регистрируется от обширных областей коры головного мозга.
В остром опыте перерезка стволовой части мозга на уровне среднего мозга и, таким образом, разрушение восходящих путей от ретикулярной формации ствола переводят животное в сноподобное коматозное состояние (спящий мозг,
по
Бремеру)
с соответствующими изменениями характера электроэнцефалограммы.
Приведенные выше экспериментальные факты послужили основанием для заключения, что ретикулярная формация является структурой, отвечающей за состояние бодрствования, структурой, формирующей восходящую активирующую ретикулярную систему, которая поддерживает на определенном уровне возбудимость промежуточного мозга и коры больших полушарий. Согласно современным представлениям, переход коры к активному состоянию связан
| Рис. 3.23 Афферентные и эфферентные связи ретикулярной формации стволовой части мозга |
с
колебаниями количества восходящих сигналов от ретикулярной формации ствола. Количество этих сигналов зависит от поступления в ретикулярную формацию сенсорных импульсов по коллатералям специфических афферентных восходящих путей. Практически к ретикулярной формации приходит информация от всех органов чувств по коллатералям от спинно—ретикулярного пути, проприоспинальных путей, афферентных черепных нервов, от таламуса и гипоталамуса, от моторных и сенсорных областей коры (рис. 3.23).
Микроэлектродная регистрация электрической активности нейронов ретикулярной формации показала, что большинство из них являются полисенсорными,
т. е. отвечают на раздражение различных модальностей (световых, звуковых, тактильных и т. д.). Ретикулярные нейроны имеют большие рецептивные поля, большой скрытый период и слабую воспроизводимость реакции. Эти свойства противоположны свойствам нейронов специфических ядер и позволяют отнести ретикулярные нейроны к категории
неспецифических.
Равным образом восходящие пути ретикулярной формации в отличие от классических специфических чувствительных проекций получили название
неспецифических проекций.
Восходящие влияния ретикулярной формации высокочувствительны к действию различных фармакологических анестезирующих препаратов и так называемых успокаивающих средств (аминазин, серпазил, резерпин и др.).
Использование электрофизиологических и гистофлюоресцентных методов позволило установить медиаторную специфичность нейронов ретикулярной формации продолговатого мозга, моста и среднего мозга. Так, например, была изучена локализация монаминовых нейронов, которые начинают флюоресцировать после обработки микросрезов стволовой части мозга в парах формальдегида. Оказалось, что самая значительная группа адренергических
клеток находится в одной из областей моста, называемой «голубоватое (место) пятно». Аксоны нейронов этой области проходят в среднем переднемозговом пучке и имеют терминальные окончания в различных отделах головного мозга: мозжечке, гипоталамусе, лимбической системе, коре больших полушарий.
Тела серотонинергических
нейронов расположены преимущественно в срединной области мозгового ствола — в составе дорсального и медиального ядер шва продолговатого мозга, среднего мозга и моста. Восходящие аксоны этих нейронов идут в гипоталамус, базальные ганглии, лимбическую систему, кору больших полушарий.
В настоящее время накоплен значительный фактический материал, свидетельствующий о разнонаправленности влияний этих двух моноаминергических систем на возбудимость различных отделов головного мозга. Серотониновая система индуцирует сон и имеет непосредственное отношение к регуляции его медленноволновой фазы. Недостаток серотонина в мозгу вызывает вынужденное бодрствование. Катехоламиновая система голубоватого места, напротив стимулирует пробуждение и играет ведущую роль в регуляции парадоксальной фазы сна. При повреждении голубоватого пятна и, соответственно, недостатке норадреналина животные спят намного дольше, нежели обычно.
Реципрокность моноаминергических систем проявляется не только в процессах регуляции сна, но и в воздействии на эмоциональную сферу организма. Например, повышение концентрации мозгового норадреналина влечет за собой усиление стрессорного состояния животных, которое может быть снято (купировано) введением исходного продукта синтеза серотонина — 5—окситриптофана. Равным образом разрушение ядер шва, где синтезируется серотонин, усиливает агрессивность подопытных крыс.
Разнонаправленные влияния моноаминергических систем мозга имеют место и в отношении регуляции двигательной активности (серотониновая система ее снижает), и в процессах формирования сложных форм поведения (выработка условных рефлексов). Это свидетельствует о модулирующей функции данных систем, которые имеют обширные связи с различными отделами головного мозга.
Другие медиаторные системы в составе ретикулярной формации стволовой части мозга представлены холинергическими нейронами, а также сопутствующими им глицинергическими клетками, которые чаще всего выполняют функции тормозных интернейронов.
Следует отметить, что в составе восходящих путей ретикулярной формации имеются активирующие и дезактивирующие группы. По данным Дж. Моруцци, раздражение некоторых участков ретикулярной формации заднего мозга может вызвать у животного глубокий сон и появление α—ритма в электроэнцефалограмме. Вероятно, реципрокный принцип организации восходящих и нисходящих проекций является общим для всей системы ретикулярной формации.
Каковы функции продолговатого мозга
Функции, выполняемые продолговатым мозгом человека, важны для организма. Без этого отдела головного мозга человек не сможет жить, так как он контролирует жизненно-важные органы тела – легкие и сердечную мышцу. Поэтому при травмировании этого мозгового отдела ЦНС наступает моментальная смерть. За всю эту деятельность отвечают ядра – скопления серого вещества.
Таблица функций ядер продолговатого мозга:
| Название структуры | Задачи, которые она выполняет |
| Ядра V-XII пары черепно-мозговых нервов | Сенсорные, двигательные, вегетативные функции заднего мозга |
| Серое вещество тонкого и клиновидного пучка | Отвечают за тактильную и проприоцептивную восприимчивость |
| Ядра оливы | Являются частью центра равновесия |
| Дорсальное ядро трапециевидного отдела | Участвуют в анализе слуховой информации |
| Серое вещество ретикулярной формации | В зависимости от ситуации активируют или тормозят нейроны спинного мозга и зоны коры головного отдела ЦНС, образуют вегетативные центы |
| Голубое пятно | Структуры способны выбрасывать специфичный гормон, который влияет на возбудимость отдельных зон коры церебрума |
Основные функции
Таким образом, функции продолговатого мозга человека подразделяются на несколько категорий:
- Осуществление и контроль безусловных рефлексов (рефлекторная функция);
- Моторика защитных и ориентировочных действий (сенсорная функция);
- Автоматизм процесса дыхания и кровообращения (интегративная функция);
- Поддержание баланса положения туловища и мышечного тонуса (проводниковая функция).
Сенсорные
Сенсорный отдел контролирует и обрабатывает прием входящих импульсов, которые идут к нему от рецепторов со всего тела человека. Эти восприимчивые структуры состоят из 2 единиц:
- Сенсоэпителиальных клеток;
- Аксонов афферентных нейронов.
Там, например, происходит обработка импульсов от дыхательного центра – оцениваются показатели крови и структура легочной ткани. По итогам этих данных в дальнейшем происходит насыщение крови кислородом и процессы метаболизма в организме. Сенсорный отдел продолговатого мозга получает и отдает в дальнейшую обработку импульсы от органов пищеварения, слуха, зрения, тактильных точек.
Продолговатый мозг воспринимает тактильную информацию с помощью следующих структур:
- При активации рецепторов кожи лица происходит возбуждение нейронов тройничного нерва;
- Рецепторов вкуса – языкоглоточного нерва;
- Звуковых рецепторов – улиткового нерва;
- При нарушении устойчивости – оливы.
Исходя из полученных данных, следует дальнейшая реакция организма, которая выражается в реализации безусловных рефлекторных реакций. Их также осуществляет продолговатый отдел ЦНС. При недостаточном насыщении крови кислородом, человек начинает глубоко дышать и искать источник свежего воздуха, тем самым активируя моторную функцию отдела.
Проводниковые
Продолговатый отдел – связующее звено спинного и конечного отдела ЦНС. Через него проходят афферентные и эфферентные тракты: кортикоспинальный, спинно-таламический, руброспинальный.
С помощью них передается информация в высшие структуры церебрума и возвращается обратно к органам в виде импульсов. Здесь начинаются оливоспинальный, вестибулоспинальный и ретикулоспинальный тракты. Через них стабилизируется напряжение мышц и координация движений рук и ног.
В продолговатом мозге заканчиваются корковоретикулярные тракты из коры, а также постилающиеся вверх волокна проприоцептивной восприимчивости из спинного мозга. Эта особенность позволяет чувствовать части собственного тела относительно друг друга в пространстве.
Интрегративная
Интрегративная функция продолговатого мозга заключается в осуществлении реакций организма несвязанных с рефлекторной деятельностью. Их появление связывают с работой нейронов конечного отдела, которые запрограммированы на исполнение регуляторных процессов в определенном порядке. Их осуществление идет через другие центры НС или напрямую.
Движения глазных яблок при наклоне головы реализуется на основе совместной деятельности ядер вестибулярной и глазодвигательной систем мозга. Часть нервных клеток ретикулярной субстанции продолговатого мозга осуществляет контроль над тонусом и координацией других нервных центров церебрума.
Функции
Продолговатый мозг отвечает за следующие основные функции:
Сенсорные функции
От сенсорных рецепторов поступают афферентные сигналы к ядрам нейронов продолговатого мозга. Затем проводится анализ сигналов:
- Системы дыхания — газовый состав крови, pH, текущее состояние растяжении легочной ткани
- Кровообращения — работа сердца, артериальное давление
- сигналы от пищеварительной системы
Результатом анализа является последующая реакция в виде рефлекторной регуляции, которую реализуют центры продолговатого мозга.
К примеру, накопление С02 в крови и снижение O2 является причинной для следующих поведенческих реакций, отрицательные эмоции, удушье и тд. которые заставляют человека искать чистый воздух.
Проводниковая функция
Эта функция заключается в проведении нервных импульсов и в самом продолговатом мозге и к нейронам других отделов мозга. Афферентные нервные импульсы приходят по одноименным волокнам 8-12пар черепных нервов к продолговатому мозгу. Также через этот отдел проходят проводящие пути от спинного мозга к мозжечку, таламусу и ядрам ствола.
Рефлекторные функции
К основным рефлекторным функциям можно отнести регуляцию тонуса мышц, защитные рефлексы и регуляцию жизненно важных функций.
Проводящие пути начинаются в ядрах ствола мозга, кроме кортикоспинального пути. Пути заканчиваются в у-мотонейронах и интернейронах спинного мозга. С помощью таких нейронов можно контролировать состояние мышц антагонистов, антагонистов и синергистов. Позволяет подключать к простым движением дополнительных мышц.
- Выпрямительные рефлексы — восстанавливает положение тела и головы. Рефлексы работают с помощью вестибулярного аппарата, рецепторов растяжения мышц. Иногда работа рефлексов настолько быстра, что мы со временем осознаем их действие. К примеру, действие мышц при соскальзывании.
- Позные рефлексы — нужны для поддержания определенной позы тела в пространстве, включая нужные мышцы
- Лабиринтные рефлексы — обеспечивают постоянное положение головы. Делятся на тонические и физическими. Физические — поддерживают позу головы при нарушении равновесия. Тонические — поддерживают позу головы на протяжении длительного времени за счет распределения контроля в разных мышечных группах
Защитные рефлексы:
- Рефлекс чиханья — за счет химического или механического раздражения рецепторов слизистой оболочки полости носа происходит форсированных выдох воздуха через нос и рот. Данный рефлекс делят на 2 фазы: респираторную и назальную. Назальная фаза — возникает при воздействии на обонятельных и решеточных нервов. Затем афферентные и эфферентные сигналы находят в «центры чиханья» по проводящим путям. Респираторная фаза — наступает при принятии сигнала в ядрах центра чиханья и накапливается критическая масса сигналов, чтобы послать сигнал к дыхательным и моторным центрам. Центр чиханья находится в продолговатом мозге на вентромедиальной границе нисходящего тракта и ядра тройничного нерва
- Рвота — опорожнение желудка (и в тяжелых случаях кишечника) через пищевод и ротовую полость.
- Глотание — сложный акт, в котором принимают участие мышцы глотки, полости рта и пищевода
- Моргание — при раздражении роговицы глаза и его конъюнктивы